心理学におけるフロンティア
はじめに
あくびは、口の開口部、深いインスピレーション、短い無呼吸、および多かれ少なかれ遅い呼気の不随意のシーケンスである(Baenninger,1997;Walusinski and Deputte,2004;Guggisberg et al.,2010;Krestel et al., 2018). 引き出されたとき、あくびは完全に抑制することはできません。 したがって、それはステレオタイプまたは反射的なパターンとして定義されている(Lehmann、1979;Provine、1986)。, ホモ-サピエンスにおいては、酸素処理(低酸素症によって引き起こされる呼吸機能)、ストレス関連行動(覚醒によって引き起こされる)、または体温調節(温熱療法によって引き起こされる;Guggisberg et al.,2010;Massen et al.,2014;ギャラップとギャラップ,2019). 生理学的応答であるため、あくびは、時刻(Giganti and Zilli、2011)または頭蓋内/脳の温度(Gallup and Eldakar、2013)などの内的および外的要因によって影響を受ける可能性があります。,あくびは、自己指示および/または他者に表示することができる(Moyaho et al.,2017;Palagi et al., 2019). ヒトおよび非ヒト霊長類では、種に応じて、あくびが他の人に示されると、脅威を伝達することができる(Troisi et al.、1990;Deputte、1994)および/または生理学的および行動的変化(Provine et al.,1987;Leone et al.,2015;Zannella et al., 2015). ヒトにおいて、あくびは、実際の社会的存在によって阻害され、仮想的社会的存在によって阻害され得るため、社会的に変調された応答である(Gallup et al.,、2019)そして、あくびは、伝染性のあくびとして知られている現象の結果として、他の誰かのあくびによって引き起こされる可能性があるため(Provine、1989、2005)。 あくび伝染は、あくびが聞こえるが見えない場合でも誘発され得る(Arnott et al.,2009;Massen et al., 2015).
ヒトにおいて、それらの系統発生的に最も近い類人猿種(チンパンジー:Pan paniscus;bonobos:Pan troglodytes)およびアフリカザルTheropithecus gelada、伝染性のあくびは存在するだけでなく(Provine、1986;Palagi et al.,2009;Tan et al.,2017;しかし、参照:Amici et al.,,2014)しかし、特定のカテゴリーの個人が関与しているときにあくび応答が最も高いため、社会的に変調される(例えば、親族、グループメンバー、支配人;Palagi et al.,2009;Campbell and de Waal,2011,2014;Norscia and Palagi,2011;Demuru and Palagi,2012;Massen et al., 2012)., 伝染性あくびにおけるこの社会的非対称性を説明するために二つの主な議論が提示されており、これらは二つの主要な仮説に分類されている:伝染性あくびを感情移動に結びつける感情バイアス仮説(EBH)と、伝染性あくびをトップダウン注意プロセスの違いの影響を受ける運動応答とみなす注意バイアス仮説(ABH)。, 2020).,
EBHは、あくび感染率で観察される社会的非対称性は、個人間の異なる社会的結合、感情的な結合の代理人の違いを反映していると予測している。 この仮説は、あくびの伝染率が共感的傾向に従うという証拠(sensu Preston and de Waal、2002)によって支持され、最も強い感情的結合を共有する個人間で最も高い。 具体的には、Norscia and Palagi(2011)は、ヒトでは、知人や見知らぬ人に対する応答よりも親族や友人に対する応答が最も大きいことを発見しました。, 大人のチンパンジーでは、あくびの伝染は、グループ外のメンバーに比べてグループ内の間で高く(Campbell and de Waal、2011)、bonobosでは、あくびの伝染率は、互いにより多くの関係を持つ個人間で最も大きい(Demuru and Palagi、2012)。 ヒトとボノボの両方を含む比較調査において、Palagi et al. (2014)は、あくびの伝染率は、被験者が属する種よりも個人間の関係の質によって影響されることを見出した。, さらに、ヒトでは、他人の感情を識別する能力が高まり、そのような能力が低下する老齢とともに低下すると、あくび伝染は年齢とともに増加する(Wiggers and van Lieshout、1985;Anderson and Meno、2003;Saxe et al.,2004;Singer,2006;Millen and Anderson,2011;Bartholomew and Cirulli,2014). あくびの伝染率は、チンパンジーでも幼児期から成人期にかけて増加する(Madsen and Persson、2013)。ABHは、あくび伝染において観察される社会的非対称性は、社会的、視覚的注意の違いによる可能性があると予測している(Massen and Gallup、2017)。, 特に、伝染性のあくびの最高レベルは、チンパンジーまたはボノボで起こるように、ヒトおよびゲラダ、またはドミナントで起こるように、よく知られた被験者のような観察者により関連性のある個体に支払われる余分なトップダウン、選択的な視覚的注意によるものである(Yoon and Tennie、2010;Massen et al.,2012;Massen and Gallup,2017). Massen and Gallup(2017)によると、ABHは、馴染みのない被験者および/またはグループ内の被験者の顔の異なる視覚検出および視覚知覚符号化に関する既存の証拠によってバックアップされるであろう(例えば、,,Buttle and Raymond,2003;Ganel and Goshen-Gottstein,2004;Jackson and Raymond,2006;Michel et al., 2006).
本研究では、ヒトの自然環境下で9年以上にわたって収集されたあくびに関するデータを分析し、被験者が放出するあくびは聞こえるが、潜在的な応答者(聴覚あくび)によって見ることができない場合を外挿した。 あくび刺激の視覚的手掛かりが検出できなかった症例のみを考慮して,あくび感染率で以前に観察された社会的非対称性が持続するかどうかを検証した。, 特に、我々は上記の二つの仮説(EBHとABH)から導かれた以下の代替予測をテストしました。
予測1a:EBHによると、あくびの伝染率は、特定の個人に特に支払われる異なるトップダウン、選択的な視覚的注意によってではなく、個々の社会的結合-感情的な結合のプロキシ—の強さによって影響されます。, この仮説が支持されている場合、我々はあくび刺激の視覚的手がかりが除外され、聴覚伝染性あくびの率が弱く結合した個人と比較して強く結合した間で高くなるときにも社会的バイアスを観察することを期待している。
予測1b:ABHによると、弱く結合した個体と比較して強く結合した個体間のあくび伝染のレベルが高いほど、個人が家族や友人など、それらに関連する個人に支払う最も近いトップダウン、選択的な視覚的注意にリンクされるであろう。, この仮説が支持されている場合、あくび伝染において観察される社会的バイアスは、潜在的な応答者が視覚的手がかりに出席できないため、聴覚あくびだけが考慮されるときに消えるはずである。
材料と方法
データ収集と運用定義
この研究では、潜在的な応答者(以下:聴覚あくび)によってのみ聞くことができる被験者によって発 エミッタと潜在的なレスポンダは、≤5mの範囲でなければなりませんでした。, 294例の聴覚あくびは、2001年の9年間にわたって収集されたあくび発作の合計のデータセットから、2010年から2019年までのすべての発生サンプリング法(Altmann、1974)を用いて外挿された。 具体的には、聴覚あくびは、2010年から2019年05月30日から02月30日まで、18歳から77歳までのヒト白人の被験者について、職場、食事、社会的会合などの日常的な活動中に収集された。, 被験者は観察されていることに気づいておらず、明らかな外部の不安源がない状態である。 聴覚あくびデータベースには、193のあくび潜在的な応答者ダイアドが含まれていました。 状況に応じて、情報は英数字コードを介して記録され、ノート化されず、計算シートに直接入力され、携帯電話で入力されたり、紙に書かれたりしてから、計算シートに入力され、その後の精緻化のために入力されました。 年齢や人との関係などの基本的な情報は著者に知られていました。, 潜在的なレスポンダーは、トリガーに対して頭を180°回転させたとき、または物理的な視力遮断障害物が存在し、潜在的なレスポンダーがトリガーの顔と体を見ることを妨げているときに、非視力状態のようにコード化された。 トリガーおよび対応が完全に消えることはありません離(例えば、二つの別々の部屋に閉扉化)します。, 0=以前に会ったことのない見知らぬ人、1=第三の外部要素に基づいて間接的な関係を独占的に共有した知人、すなわち仕事義務(同僚)または共通の友人(友人の友人)、2=友人、第三の外部要素に排他的に関連しない直接的な関係を共有する非関連の個人、3=通常のパートナーおよび親族(r≥0.25)。, 以前の文献では、他の誰かのあくび(トリガーのあくび)を見た後、5分以内にあくび応答を引き出すことができると報告されています(Provine、1986)、最初の分で最大(Provine、2005;Palagi et al., 2014). 文献はまた、四分目から自己相関の最も高い確率があることを報告している(t0で被験者によって行われるあくびの存在は、Xが時間の増加単位であるt(0+X)で同じ被験者によって別のあくびを持つ確率を増加させることを意味する;Kapitány and Nielsen,2017)。, したがって、トリガーによって放出されたあくびから3分の時間ウィンドウ内で発生するあくび応答を考慮しました。 自己相関バイアスをさらに減らすために、トリガーによって放出される一連のあくび(3分の時間ウィンドウで放出されるより多くのあくび)の場合、最後のあくびの知覚の後に実行される最初のあくびのみを応答として登録しました。 私たちは、あくびイベントの前の5分で他の被験者があくびをしていなかったときに、あくびを”自発的”としてコード化しました。, 1=存在、0=不在、社会的結合は上記で定義された四つのレベルで入力された(0=見知らぬ人、1=知人、2=友人、3=親族)、トリガーとオブザーバーの性別はM=男性、F=女性、トリガーとレスポンダーの年齢クラスはyo=若者(18-24歳)、ad=大人(25-64歳)、se=シニア(65歳以上)(カナダ統計局、2009年)、タイムスロットは1=05:30-09のようにコード化された。:00am;2=09:01am–12:30pm;3=12:31-16:00pm;4=16:01-19:30pm;5=19:31-23:00pm;6=23:01-02:30(giganti and zilli,2011)., データベース(補足データシートを参照)には、84人の男性、69人の女性、16人の若者(yo)、122人の大人(ad)、および15人の高齢者(se)が含まれていました。, あくび感染の発生が要因bond(0=見知らぬ人;1=知人;2=友人;3=親族)、トリガーの性別(Trigger_sex)、レスポンダーの性別(Responder_sex)、トリガーの年齢クラス(Trigger_ageclass)、レスポンダーの年齢クラス(Responder_ageclass)、およびタイムスロット(1から6)によって影響されたかどうかをテストするために、一般化線形混合モデル(GLMM)を使用した。レスポンダー(応答者)は変量効果として識別します。 R(Rコアチーム、2018;バージョン3.5)にモデルを装着しました。,1)Rパッケージlme4の機能lmerを使用する(Bates et al., 2015). 変量効果のみを含むヌルモデルと比較することにより、完全モデルの重要性を確立しました(Forstmeier and Schielzeth、2011)。 この有意性をテストするために尤度比検定(Dobson、2002)を使用しました(引数”Chisq”を使用したANOVA)。 我々は、R関数”drop1″を使用して、完全モデルとヌルモデルとの間の尤度比検定に基づいて個々の予測変数のp値を計算した(Barr et al., 2013). 応答変数がバイナリであるため、二項誤差分布を使用しました。, トリガーとレスポンダーの性別または年齢クラス間の相互作用が有意であるかどうかをテストしたが、そうでなかったため、モデルには含めなかった。 我々は、多重対照パッケージ(multcomp)を使用して、Tukey検定を用いて各結合レベルに対するすべての対比較を行った(Bretz et al., 2010). Bonferroni調整p値,推定値(Est),標準誤差(SE)およびz値を報告した。
結果

図1., 音響あくび伝染の平均発生に対するトリガーとレスポンダー(X軸)間の社会的結合の影響の折れ線プロット(Y軸)。 友人と親族は、見知らぬ人や知人よりも有意に高いあくび感染頻度を示しています(Tukeyテスト:友人と知人p<0.001;親族と見知らぬ人p<0.001;友人と知人p<0.001;親族と知人p<0.001;その他の組み合わせ、ns)。,
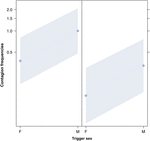
図2. 音響あくび伝染の平均発生に対するトリガーの性別(X軸)の影響の折れ線プロット(Y軸)。 応答者が女性(右)および男性(左)である場合の音響のあくびの伝染に対する制動機の性の効果。 男性のあくびは、応答者の性別にかかわらず、女性のものよりも多くのあくびを引き出す(GLMMの結果、p=0.022)。 ポイントは、モデルによって予測された値に基づいて、応答変数が独立変数”トリガーの性別”に及ぼす影響を表します。, 色付きのバンドは、95%信頼区間を示します。
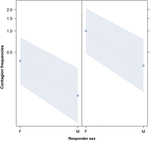
図3. 音響のあくびの伝染の平均の発生に対する応答者(X軸)の性の効果の折れ線プロット(Y軸)。 引き金が女性(右)または男性(左)の場合の音響のあくびの伝染に対する応答機の性の効果。 女性はトリガーの性別にかかわらず男性よりも有意に多く反応する(GLMMの結果、p=0.021)。, ポイントは、モデルによって予測された値に基づいて、応答変数が独立変数”応答者の性別”に及ぼす影響を表します。 色付きのバンドは、95%信頼区間を示します。
Discussion
この研究では、あくび感染が個人間の社会的結合によって大きく影響されることを初めて示しています表1トリガー刺激が聴覚的なあくびである場合であっても、私たちは聞くことができるが見られない発声されたあくび(視覚的手が検出できない、聴覚の手が検出できない)と定義した。, 特に、聴覚伝染性あくびは、見知らぬ人や知人の間よりも親族と友人の間で有意に頻繁である(図1)。 この知見は、EBHに基づく予測1aを支持し、ABHに基づく予測1bを支持しておらず、ヒトのトップダウンでは、選択的視覚的注意があくび感染率で観察される社会的非対称性の主な要因ではないという結論につながった(Norscia and Palagi,2011;Norscia et al.、2016年6月)。, また、トリガーの性別とレシーバーの性別は、あくびの伝染率に大きな影響を与え、男性はトリガーとして他の人によって女性よりも頻繁に反応し(図2)、女性は男性よりも他の人のあくびに頻繁に反応する(図3)。
Bartholomew and Cirulli(2014)とは対照的に、聴覚あくびに関するデータベースが成人(25-64歳)の強い有病率を有していたため、あくび感染に年齢効果は見つかりませんでした。, 男性と比較した女性の聴覚あくび伝染の最高レベルは、Norscia et al.によってあくび伝染の影響を受けやすいヒトの自然主義的条件で観察されたジェンダーバイアスを確認している。 (2016a、b)潜在的な応答者が見ることができるあくびを含むより大きなデータセットを検討する場合(あくび感覚モダリティ—視覚、聴覚、またはその両方—応答, ジェンダーバイアスは、視覚的な手がかりを含む制御された設定で得られた以前の結果と部分的に一致しています(Chan and Tseng、2017;ただし、Norscia and Palagi、2011;Bartholomew and Cirulli、2014)。 女性におけるあくび感染の高度は、感情的伝染を知らせる可能性があるという仮説が立てられている(Norscia et al.、2016a)、男性と比較して女性の出産に関連するより高い共感能力に照らして(Christov—Moore et al., 2014)., しかし、この問題は、人間社会全体の文化的差異が異なる方法で社会的結合ダイナミクスを形成する可能性があるため、まだ議論されている。 したがって、知識のこの段階では、文化的要因、個人間の関係の質、およびあくび伝染の分布におけるジェンダーの影響を解くことが複雑になります。 我々の結果はまた、男性がトリガーとして女性よりも優れていることを示しており、これに対する最も節約的な仮説は、男性の発声がしばしば背景雑音に, 確かに、声の性別の認識は、主に女性の声よりも男性のオクターブだけ平均して低い基本周波数に依存し、低い周波数の発声は高周波のものよりもさらに進んでいる(Marten and Marler、1977;Latinus and Taylor、2012)。 しかし、我々の知る限り、あくびの可聴性における可能なジェンダーバイアスに対処する特定の研究はなく、したがって、この憶測を検証するためには、制御,
この研究では、カテゴリー(家族や友人、見知らぬ人、知人)間のあくび感染率の違いは、トップダウンの選択的視覚的注意の違いによって説明できないこ この発見は以前の文献に沿ったものです。 伝染性のあくびは、トップダウン関連領域よりも向き付け-ボトムアップネットワークに関連する脳領域を含むように見える。 さらに、あくび伝染は、信号中に存在する感覚的手がかり(聴覚、視覚、または視聴覚)にも敏感ではない(Arnott et al.,、2009;NorsciaとPalagi、2011)も誘発刺激の視覚的視点の影響を受けない(90°、180°、および270°の向きのあくびは、正面、0°あくびとしてあくび応答を引き起こすことができる;Provine、1989、1996)。 Chan and Tseng(2017)は、あくびをそのように検出する能力(知覚検出感度)は、刺激放出顔の目への凝視の持続時間に関連していたが、目の凝視パターンは伝染性のあくびを調節することができなかったことを見出した。, チンパンジーでは、レスポンダーがグループ外のチンパンジーのビデオをより長く見たとしても、伝染性のあくび頻度は異なるグループの個体よりも同じグループの間で最も高かった(Campbell and de Waal、2011)。 視覚的選択的注意が特定の方向(見知らぬ人よりも家族に反応する被験者)にあくびの伝染率を偏らせることができるという議論は、ヒトおよび非ヒト霊長類における社会的注意の特定のパターンがないことによっても損なわれる。 アイトラッキング(視聴時間を測定するために適用される)を介して、および未知の顔を実験対象に示すことによって、Méary et al., (2014)では、ヒトは自分の人種の顔に偏っているのに対し、アカゲザルの注意は同じ種の顔よりも新しい顔によってより引き付けられていることが観察された。 川上ほか (2014)は、ヒト被験者が民族内グループメンバーの目と民族外グループメンバーの鼻と口にもっと注意を払っていることを観察した。 同じ研究では、視覚的注意が対象レースに依存しないことも明らかになった。 実験対象がスクリーンを見つめた時間を測定することによって、Whitehouse et al., (2016)は、バーバリーマカクが見知らぬ人よりも非知らぬ人のビデオを傷つけることにもっと注意を払ったことを観察したが、非知らぬ人の中では、マカクが弱い社会的結合を共有している個人に最も注意を払ったことにも注意を払ったことを指摘した。 一目率を測定することによって、SchinoとSciarretta(2016)は、マンドリルは非親族よりも自分の親族に多く見えるが、下位グループの仲間よりも優性に多く見えること したがって、これらの研究(ABHを支持するために使用)は、選択的注意の単一のパターンを記述しない。, 考慮すべきさらに重要な点は、ABHをサポートするために使用された研究のほとんどで採用された親しみやすさとグループメンバーシップの定義です(Massen and Gallup、2017)。 これらの研究は、慣れ親しんだ/グループ内の被験者の顔のより良い視覚検出と視覚知覚エンコーディングを示したが、個人間の個人的な関係に基づいてではなく、親しみやすさとグループメンバーシップを定義した。 代わりに、親しみやグループメンバーシップは、間接的な知識の地面に定義されました(例えば, 例えば、Buttle and Raymond,2003;Ganel and Goshen-Gottstein,2004;Jackson and Raymond,2006;Michel et al., 2006). この定義は、これらの研究の目的のためには問題ありませんが、実際の関係に基づく実際の社会的結合が現象、この場合はあくび伝染に及ぼす影響についての代替説明を提案するために結果が使用されている場合、それほど問題はありません。 例えば、Michel et al., (2006)は、白人とアジアの被験者が同じ人種の顔をよりよく認識できることを観察したが、この違いは白人の間で約一年間住んでいたアジアの被験者には 別の議論のポイントは、視覚的注意の変化を頻繁に示す自閉症スペクトラム障害(ASD)の子供におけるあくび伝染の存在に関するものである(Richard and Lajiness-O’Neill、2015)。 ASD小児では、あくび伝染は存在しない可能性がある(Senju et al.、2007)、障害(Helt et al.,,2010)、または被験者が実験試験中にビデオ刺激に注意を向けるように誘導されたときに典型的に発達している子供に類似している(Usui et al., 2013). 最近の研究では、Mariscal et al. (2019)は、ASD小児におけるあくび伝染は、親および社会的愛着に関与するホルモンであるオキシトシンの血中濃度と正の関連があることを見出した(Decety et al.、2016)およびASD小児におけるあくび伝染は、異なる研究コホート間の可変平均オキシトシン濃度に関連している可能性があると仮定した(Mariscal et al., 2019)., この発見は、あくびの伝染率を社会的結合に結びつけるというEBH仮説に沿ったものであり、感情的な結合を反映する可能性がある。
結論
私たちの研究は、あくび伝染における社会的非対称性の根底にあるメカニズムについての議論に加えて、批判的なレビューについては、Adriaense et al.,2020;Palagi et al.,2020)、あくびの伝染は、おそらくトップダウンの選択的な注意ではなく、ボトムアップに関連していることを示すことによって。, ボトムアップの注意は、主に誘発刺激の感覚知覚によって導かれるのに対し、トップダウンの選択的注意は、特定の項目が内部的に選択され、焦点を当てたり検査されたりする自発的で持続的なプロセスである(Katsuki and Constantinidis、2014)。 この点で、トリガーによって放出される音響刺激(聴覚あくび)が聞こえ、受信機がトリガーに任意の視覚的注意を払っていなかったにもかかわらず、受信機であくび応答を引き出すことができた。, さらに、あくび応答率は社会的に変調され、聴覚あくび伝染は互いに最も強く結合していた個人で最も高かった。 したがって、トップダウン選択的注意は、ボトムアップ注意プロセスに関連する刺激駆動現象であるように見えるあくび伝染で観察される社会的非対称性の主なドライバーではありません。 他の形態の注意または注意深い段階があくびの伝染に影響を与えることができるかどうか、そしてどのような方法で理解するためにさらなる調査,
データ可用性ステートメント
この研究に使用されるデータセットは、補足資料に記載されています。
倫理声明
人間の参加者を含む研究は、Comitato di Bioetica d’Atteneo(大学生命倫理委員会)—トリノ大学(ref. 番号451945)。 同意書への参加については不要であったためこの研究に従って各国の国内法や制度的条件です。
著者の貢献
INとEPは、データ収集を行い、原稿を考案し、執筆しました。, AZは、データ収集と原稿の改訂、図、および表を支援しました。 MGは統計分析を行い、原稿の関連部分を書いた。
利益相反
著者らは、この研究は、潜在的な利益相反と解釈され得る商業的または財政的関係がない場合に行われたと宣言している。
謝辞
著者らは、それぞれピサ大学とトリノ大学での研究を支援してくれたRoberto Barbuti教授とCristina Giacoma教授に感謝したいと考えています。,
Supplementary Material
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–266. doi: 10.1163/156853974×00534
PubMed Abstract | CrossRef Full Text | Google Scholar
Anderson, J. R., and Meno, P. (2003). Psychological influences on yawning in children. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J., and Cirulli, E. T. (2014)., 伝染性のあくび感受性の個人差は非常に安定しており、共感または他の既知の要因によって主に説明されていない。 PLoS One9:e91773. 土井:10.1371/ポーン0091773
PubMed Abstract|CrossRef Full Text|Google Scholar
Bretz,F.,Hothorn,T.,and Westfall,P.(2010). R.Boca Ratonを用いた多重比較,FL:Chapman&Hall.
Google Scholar
Deputte、B.L.(1994)。 霊長類におけるあくびの生態学的研究。 I., 旧世界ザル(Cercocebus albigenaとMacaca fascicularis)の二つの種における因果関係の定量分析と研究。 エソロジー98,221-245. doi:10.1111/j.1439-0310.1994.01073x
CrossRef Full Text|Google Scholar
Dobson,A.J.(2002). 一般化線形モデルの紹介、第2Edn。 ボカラトン、フロリダ州:チャップマンとホール。
Google Scholar
Katsuki,F.,And Constantinidis,C.(2014). ボトムアップとトップダウンの注意:異なるプロセスと重複する神経系。, 20509-521 doi:10.1177/1073858413514136
PubMed Abstract|CrossRef Full Text|Google Scholar
Singer,T.(2006). 共感と心の読書のニューロンの基礎と個体発生:文献のレビューと将来の研究への影響。 ニューロシティ バイオベハブ 牧師30,855-863. 土井:10.1016/j.neubiorev.2006.06.011
PubMed Abstract|CrossRef Full Text|Google Scholar
カナダ統計局(2009). 年齢カテゴリ、ライフサイクルグループ。 オタワ:カナダ統計局。,
Google Scholar