Fronteiras em Psicologia
Introdução
o Bocejo é involuntário sequência de abertura de boca, inspiração profunda, breve apnéia, e mais ou menos lenta expiração (Baenninger, 1997; Walusinski e Deputte, 2004; Guggisberg et al., 2010; Krestel et al., 2018). Quando provocado, um bocejo não pode ser totalmente suprimido. Portanto, tem sido definido como um padrão estereotipado ou reflexivo (Lehmann, 1979; Provine, 1986)., No Homo sapiens, várias hipóteses têm sido apresentadas com suporte variável para explicar mecanismos e funções de bocejo espontâneo, tais como oxigenação (função respiratória causada por hipoxia), comportamento relacionado ao estresse (causado por excitação), ou termorregulação (causada por hipertermia; Guggisberg et al., 2010; Massen et al., 2014; Gallup e Gallup, 2019). Sendo uma resposta fisiológica, o bocejo pode ser afetado por fatores internos e externos, como a hora do dia (Giganti e Zilli, 2011) ou temperatura intracraniana/cerebral (Gallup e Eldakar, 2013).,o bocejo pode ser auto-dirigido e / ou exibido para outros (Moyaho et al., 2017; Palagi et al., 2019). Em primatas humanos e não humanos, dependendo da espécie, quando o bocejo é mostrado aos outros, pode comunicar a ameaça (Troisi et al., 1990; Deputte, 1994) and / or physiological and behavioral changes (Provine et al., 1987; Leone et al., 2015; Zannella et al., 2015). Em humanos, bocejar é uma resposta socialmente modulada porque pode ser inibida pela presença social real—e não virtual – (Gallup et al.,, 2019) e porque um bocejo pode ser desencadeado pelo bocejo de outra pessoa, como resultado de um fenômeno conhecido como bocejo contagioso (Provine, 1989, 2005). O contágio do bocejo pode ser provocado mesmo que o bocejo seja ouvido, mas não visto (Arnott et al., 2009; Massen et al., 2015).em humanos, suas espécies de macacos filogeneticamente mais próximas (chimpanzés: Pan paniscus; bonobos: Pan troglodytes) e o macaco Africano Theropithecus gelada, bocejo contagioso não está apenas presente (Provine, 1986; Palagi et al., 2009; Tan et al., 2017; but see: Amici et al.,, 2014), mas também socialmente modulada porque a resposta bocejante é maior quando certas categorias de indivíduos estão envolvidos (por exemplo, kin, membros de grupo, dominantes; Palagi et al., 2009; Campbell and De Waal, 2011, 2014; Norscia and Palagi, 2011; Demuru and Palagi, 2012; Massen et al., 2012)., Dois principais argumentos que têm sido apresentadas para explicar esta assimetria social em contagiosa bocejando, e que foram agrupados em duas hipóteses principais: a Emocional Viés Hipótese (EBH), ligando o bocejo contagioso emocional de transferência, e o Viés Atencional Hipótese (ABH), que considera o bocejo contagioso como uma resposta motora que está sujeita a diferenças no top-down processos atencionais (Palagi et al., 2020).,
A EBH prevê que a assimetria social observada nas taxas de contágio bocejado reflete diferenças nas diferentes ligações sociais, um proxy da ligação emocional, entre indivíduos. Esta hipótese é apoiada por evidências de que as taxas de contágio bocejadas seguem uma tendência empática (sensu Preston e de Waal, 2002), sendo a mais alta entre os indivíduos que compartilham um vínculo emocional mais forte. Especificamente, Norscia e Palagi (2011) descobriram que em humanos as taxas de contágio são maiores em resposta a parentes e amigos do que em resposta a conhecidos e estranhos., Em chimpanzés adultos o contágio do bocejo é maior entre os membros do grupo em comparação com os membros do grupo (Campbell e de Waal, 2011) e em bonobos as taxas de contágio do bocejo são maiores entre os indivíduos que se afiliam mais uns com os outros (Demuru e Palagi, 2012). Em uma investigação comparativa, incluindo humanos e bonobos, Palagi et al. (2014) constatou que as taxas de contágio bocejadas foram afetadas pela qualidade da relação entre indivíduos mais do que pela espécie a que os sujeitos pertenciam., Além disso, nos seres humanos, o contágio do bocejo aumenta com a idade, quando a capacidade de identificar as emoções dos outros aumenta e diminui com a idade, quando essa capacidade diminui (Wiggers e van Lieshout, 1985; Anderson e Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen and Anderson, 2011; Bartholomew and Cirulli, 2014). As taxas de contágio do bocejo aumentam desde a infância até a idade adulta também em chimpanzés (Madsen e Persson, 2013).
a ABH prevê que a assimetria social observada no contágio do bocejo pode ser devido a diferenças na atenção social, visual (Massen e Gallup, 2017)., Em particular, os mais altos níveis de contagiosa, o bocejo seria devido o adicional de cima para baixo, seletiva visual a atenção prestada aos indivíduos que são mais relevantes para o observador, como familiar assuntos, como ocorre em seres humanos e geladas, ou dominantes, como ocorre em chimpanzés e bonobos (Yoon e Tennie, 2010; Massen et al., 2012; Massen and Gallup, 2017). De acordo com Massen e Gallup (2017), a ABH seria apoiada pelas evidências existentes sobre a diferente detecção visual e codificação visual perceptiva das faces de indivíduos familiares e / ou em grupo em comparação com os desconhecidos (e.g.,, Buttle and Raymond, 2003; Ganel and Goshen-Gottstein, 2004; Jackson and Raymond, 2006; Michel et al., 2006).
neste estudo, analisamos dados sobre bocejos coletados ao longo de 9 anos em seres humanos em suas configurações naturais e extrapolamos os casos em que o bocejo emitido por um sujeito poderia ser ouvido, mas não visto por um potencial respondedor (bocejo auditivo). Ao considerar apenas os casos em que a deixa visual do estímulo bocejante não era detectável, verificamos se a assimetria social anteriormente observada nas taxas de contágio bocejado persistia ou não., Em particular, testamos as seguintes previsões alternativas derivadas das duas hipóteses apresentadas acima (EBH e ABH).
Previsão 1a: de acordo com o EBH, as taxas de contágio bocejante são influenciadas pela força do vínculo social inter-individual—um proxy do vínculo emocional—por si só e não por uma atenção visual selectiva de cima para baixo, dada a certos indivíduos em particular., Se esta hipótese é apoiada, esperamos observar o viés social também quando a deixa visual do estímulo bocejante é excluída e as taxas de bocejo auditivo contagioso a ser maior entre fortemente ligados em comparação com indivíduos fracamente ligados.
Previsão 1b: de acordo com a ABH, os níveis mais elevados de contágio do bocejo entre indivíduos fortemente ligados em comparação com indivíduos fracamente ligados estariam ligados à atenção visual mais próxima, top-down, seletiva que os indivíduos prestam aos indivíduos que são relevantes para eles, por exemplo, família e amigos., Se esta hipótese é suportada, o viés social observado no contágio bocejado deve desaparecer quando somente bocejos auditivos são considerados porque a deixa visual não pode ser atendida pelo potencial respondedor.materiais e métodos
recolha de dados e definições operacionais
para este estudo, considerámos os bocejos vocalizados emitidos por um sujeito que só podia ser ouvido—mas não visto—por um potencial respondedor (a seguir: bocejos auditivos). O emissor e os potenciais respondedores tinham de estar num intervalo ≤5 m., Vocalizado bocejos envolver o uso de pregas vocais e os bocejos que apenas envolvida forte inspiração/expiração não foram considerados como vocalizado; 294 casos de auditivo bocejos foram extrapolados a partir de um conjunto de dados de um total de 2001 bocejando acessos coletados ao longo de 9 anos—a partir de 2010 a 2019—usando a todas as ocorrências método de amostragem (Altmann, 1974). Especificamente, bocejos auditivos foram coletados de novembro de 2010 a maio de 2019, das 05h30 às 02h30, em indivíduos humanos caucasianos, com idades entre 18 e 77 anos, durante suas atividades rotineiras, por exemplo, em locais de trabalho, durante refeições, durante reuniões sociais, etc.,, sem que os indivíduos tenham conhecimento de ser observados e na ausência de qualquer fonte externa evidente de ansiedade. A base de dados de bocejos auditivos incluía 193 diads com potencial de resposta. Dependendo da situação, a informação foi registrada, não anotada, através de códigos alfanuméricos e entrou diretamente em folhas de cálculo, digitadas em telefones celulares ou escritas em papel, e depois entrou em folhas de cálculo para posterior elaboração. Informação básica, como a idade e a relação entre as pessoas era conhecida pelos autores., Os potenciais respondedores foram codificados como no estado de não visão quando a sua cabeça foi rodada 180° em relação ao gatilho ou quando estava presente um obstáculo físico e bloqueador de visão que impedia o potencial respondedor de ver a face e o corpo do gatilho. O gatilho e o respondedor nunca foram completamente isolados (por exemplo, em dois quartos separados com portas fechadas) um do outro., A proximidade social foram coletados em quatro níveis: 0 = estranhos, que nunca tinha encontrado antes; 1 = conhecidos, que apenas compartilham de uma relação indireta com base em um terceiro elemento externo, que é o trabalho dever (colegas) ou amigos em comum, (amigos dos amigos); 2 = amigos, não relacionados a indivíduos que partilham uma relação direta, ainda que não exclusivamente relacionadas a um terceiro elemento externo; 3 = regular, parceiros e parentes (r ≥ 0.25)., A literatura anterior relata que as respostas bocejadas podem ser elicitadas dentro de 5 minutos depois de observar o bocejo de outra pessoa (o bocejo do gatilho) (Provine, 1986), com um máximo no primeiro minuto (Provine, 2005; Palagi et al., 2014). A literatura também relata que a partir do quarto minuto há uma maior probabilidade de autocorrelação (o que significa que a presença de um bocejo realizada por um sujeito em t0 aumenta a probabilidade de ter outro bocejo por um mesmo indivíduo em t(0+X), onde X é a unidade de aumento do tempo; Kapitány e Nielsen, 2017)., Por isso, considerámos as respostas bocejadas que ocorrem dentro de um intervalo de tempo de 3 minutos a partir do bocejo emitido pelo gatilho. Para reduzir ainda mais o viés de autocorrelação, no caso de uma cadeia de bocejos emitidos pelo gatilho (mais bocejos emitidos na janela de tempo de 3 minutos) registramos como Resposta apenas o primeiro bocejo realizado após a percepção do último bocejo. Codificámos um bocejo como “espontâneo” quando nenhum outro sujeito tinha bocejado nos 5 minutos que antecederam o bocejo., codificado como: 1 = presença, 0 = ausência; o laço social foi inserido com quatro níveis definidos acima (0 = estranhos; 1 = conhecidos; 2 = amigos; 3 = parentes); trigger e observador do sexo, foram rotulados como: M = masculino, F = feminino; a idade classes do gatilho e o respondente foram codificados como segue: yo = jovens (18-24 anos); ad = de adultos (25-64 anos); se = sênior (acima de 65 anos de idade) (Statistics Canada, 2009); os intervalos de tempo foram codificados da seguinte forma: 1 = 05:30-09:00 am; 2 = 09:01 am–12:30 pm; 3 = 12:31-16:00 pm; 4 = 16:01-19:30 pm; 5 = 19:31-23:00 pm; 6 = 23:01-02:30 (Giganti e Zilli, 2011)., A base de dados (Ver ficha complementar) incluía 84 homens, 69 mulheres, 16 jovens (yo), 122 adultos (ad) e 15 idosos (se)., Para testar se a ocorrência de contágio do bocejo foi influenciado pelos fatores bond (0 = estranhos; 1 = conhecidos; 2 = amigos; 3 = kin), sexo do gatilho (Trigger_sex), sexo do respondente (Responder_sex), a classe de idade dos trigger (Trigger_ageclass), e a classe de idade dos respondentes (Responder_ageclass), e intervalo de tempo (de 1 a 6), utilizou-se um generalizado linear misto (modelo de GLMM) que incluiu os cinco preditores como efeitos fixos e gatilhos (Trigger) e respondentes (Respondente) identidades como efeitos aleatórios. Adaptamos os modelos em R (R Core Team, 2018; Versão 3.5.,1) usando a função lmer da R-package lme4 (Bates et al., 2015). Estabelecemos o significado do modelo completo em comparação com um modelo nulo que compreende apenas os efeitos aleatórios (Forstmeier e Schielzeth, 2011). Nós usamos um teste de relação de probabilidade (Dobson, 2002) para testar este significado (ANOVA com argumento “Chisq”). Calculamos os valores p para os predictores individuais com base em testes de relação de probabilidade entre o modelo completo e o nulo usando a função R “drop1” (Barr et al., 2013). Como a variável resposta era binária, nós usamos uma distribuição de erro binomial., Testamos se a interação entre os sexos ou as classes etárias do gatilho e do respondedor eram significativas, mas como não eram, não as incluímos no modelo. Nós usamos um pacote de contraste múltiplo (multcomp) para realizar todas as comparações emparelhadas para cada nível de ligação com o teste Tukey (Bretz et al., 2010). Nós relatamos os valores p ajustados de Bonferroni, estimativa (Est), erro padrão (SE), e valores z -.
Resultados

a Figura 1., Gráfico de linha do efeito da ligação social entre o gatilho e o respondedor (eixo X) na ocorrência média de contágio acústico de bocejo (eixo Y). Amigos e parentes significativamente maior contágio do bocejo frequências de estranhos e conhecidos (Tukey, teste: os amigos versus estranhos p < 0.001; parentes versus estranhos p < 0.001; amigos versus conhecidos p < 0.001; parentes versus conhecidos p < 0.001; outras combinações, ns).,
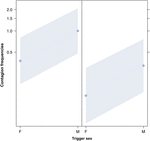
Figura 2. Gráfico de linha do efeito do sexo do gatilho (eixo dos X) na ocorrência média do contágio do bocejo acústico (eixo dos Y). Efeito do sexo desencadeador sobre o contágio acústico do bocejo quando o respondedor é uma fêmea (direita) e um macho (esquerda). Os bocejos dos machos provocam mais bocejos do que os das fêmeas, independentemente do sexo do respondedor (resultado do GLMM, p = 0,022). Os pontos representam o efeito que a variável de resposta tem sobre a variável independente “sexo do gatilho”, com base no valor previsto pelo modelo., As bandas coloridas mostram o intervalo de confiança de 95%.
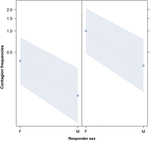
a Figura 3. Gráfico de linha do efeito do sexo do respondedor (eixo X) na ocorrência média do contágio do bocejo acústico (eixo Y). Efeito do sexo do respondedor no contágio acústico do bocejo quando o gatilho é uma fêmea (direita) ou um macho (esquerda). As fêmeas respondem significativamente mais do que os machos, independentemente do sexo do gatilho (resultado do GLMM, p = 0.021)., Os pontos representam o efeito que a variável resposta tem sobre a variável independente “sexo do respondedor”, com base no valor previsto pelo modelo. As bandas coloridas mostram o intervalo de confiança de 95%.
Discussão
Este estudo mostra pela primeira vez que o contágio do bocejo é significativamente afetado pelo laço social entre os indivíduos (Tabela 1), mesmo quando o desencadeamento de estímulos são auditivo bocejos, que definimos como vocalizado bocejos que podia ser ouvido, mas não visto (indicação visual indetectável, auditivo cue detectável)., Em particular, o bocejo auditivo contagioso é significativamente mais frequente entre parentes e amigos do que entre estranhos e conhecidos (Figura 1). Este achado suporta a previsão 1a com base na EBH e não a previsão 1b com base na ABH, levando à conclusão de que em humanos de cima para baixo, a atenção visual seletiva não pode ser o principal motor da assimetria social observada em taxas de contágio bocejadas (Norscia e Palagi, 2011; Norscia et al., 2016a)., Também o sexo do gatilho e o sexo do receptor tiveram um efeito significativo nas taxas de contágio do bocejo, com os homens—como gatilhos—sendo respondidos por outros com mais frequência do que as mulheres (Figura 2) e as mulheres respondendo mais frequentemente aos bocejos dos outros do que os homens (Figura 3).
Ao contrário de Bartolomeu e Cirulli (2014), não encontramos nenhum efeito de idade no contágio bocejado, muito provavelmente porque nosso banco de dados sobre bocejos auditivos teve uma forte prevalência de adultos (25-64 anos de idade)., Os níveis mais elevados de contágio auditivo nas mulheres em comparação com os homens confirmam o viés de gênero observado em condições naturalísticas nos seres humanos suscetíveis ao contágio bocejado por Norscia et al. (2016a, b) ao considerar um conjunto de dados maior que também incluísse bocejos que pudessem ser vistos pelo potencial respondedor (com bocejando modalidade sensorial—visão, audição, ou ambos—não afetando a resposta)., O preconceito de gênero também está em concordância parcial com resultados anteriores obtidos em configurações controladas, incluindo o visual cue (Chan e Tseng, 2017; mas veja Norscia e Palagi, 2011; Bartholomew e Cirulli, 2014). Foi colocada a hipótese de que o alto grau de contágio nas mulheres pode informar o contágio emocional (Norscia et al., 2016a), à luz das capacidades empáticas alegadamente mais elevadas-relacionadas com a maternidade—das mulheres em comparação com os homens (Christov-Moore et al., 2014)., No entanto, esta questão ainda está em debate porque as diferenças culturais entre as sociedades humanas podem moldar a dinâmica de ligação social de uma forma diferente. É, portanto, complicado, nesta fase do conhecimento, separar os fatores culturais, a qualidade das relações inter-pessoais e a influência do gênero na distribuição do contágio bocejado. Nossos resultados também mostram que os homens atuam melhor do que as mulheres como gatilhos e a hipótese mais parsimônica para isso pode ser que as vocalizações dos homens podem ser melhor ouvidas em ambientes naturais, muitas vezes caracterizados por ruídos de fundo., Na verdade, a percepção do sexo da voz baseia-se principalmente na frequência fundamental que é, em média, mais baixa por uma oitava em vozes masculinas do que femininas, com vocalizações de menor frequência viajando além das de alta frequência (Marten e Marler, 1977; Latinus e Taylor, 2012). No entanto, tanto quanto sabemos, não existe um estudo específico sobre o possível viés de gênero na audibilidade bocejada e, por isso, é necessária uma investigação adicional com ensaios experimentais em condições controladas para verificar esta especulação.,
neste estudo, também descobrimos que as diferenças nas taxas de contágio de bocejos entre categorias (família e amigos, estranhos e conhecidos) não podem ser explicadas por diferenças na atenção visual selectiva de cima para baixo. Este achado está em linha com a literatura anterior. Bocejos contagiosos parecem envolver áreas cerebrais que estão mais relacionadas com a rede orienting-bottom up do que áreas relacionadas de cima para baixo . Além disso, o contágio bocejado não é nem sensível aos sinais sensoriais presentes no sinal (auditivo, visual ou audio-visual) (Arnott et al.,, 2009; Norscia e Palagi, 2011), nem afetado pela perspectiva visual do desencadeamento de estímulo (boceja em orientações de 90°, 180° e 270° são capazes de desencadear bocejando respostas frontal, 0° bocejos; Provine, 1989, 1996). Chan e Tseng (2017) descobriram que a capacidade de detectar um bocejo como tal (sensibilidade de detecção perceptual) estava relacionada com a duração do olhar para os olhos do estímulo libertando a face, mas os padrões de olhar-olho não foram capazes de modular o bocejo contagioso., Em chimpanzés, as freqüências de bocejo contagioso foram mais altas entre o mesmo grupo do que os indivíduos de diferentes grupos, mesmo que os respondedores olhassem mais tempo para vídeos de chimpanzés fora do grupo (Campbell e de Waal, 2011). O argumento de que a atenção seletiva visual pode distorcer as taxas de contágio em uma direção específica (sujeitos respondendo mais à família do que estranhos) também é minado pela ausência de qualquer padrão específico de atenção social em primatas humanos e não humanos. Através do acompanhamento dos olhos (aplicado para medir o tempo de visualização) e mostrando rostos desconhecidos aos seus sujeitos experimentais, Méary et al., (2014) observed that humans were skewed toward own-race faces whereas rhesus macaques’ attention was more attracted by new than by same species faces. Kawakami et al. (2014) observed that human subjects paid more attention to the eyes of ethnic in-group members and to nose and mouth of ethnic out-group members. O mesmo estudo também revelou que a atenção visual não dependia da raça alvo. Medindo quanto tempo os sujeitos experimentais olharam para o ecrã, Whitehouse et al., (2016) observaram que, macacos, pois eles não prestou mais atenção para coçar vídeos de não-estranho, que estranho indivíduos, mas também ciente de que o não-estranhos, macacos pago mais atenção aos indivíduos com os quais eles compartilham de uma fraca ligação social. Ao medir as taxas de relance, Schino e Sciarretta (2016) observaram que os mandrills olhavam mais para os seus próprios parentes do que para os não-parentes, mas também mais dominantes do que para os companheiros de grupo subordinados. Por conseguinte, estes estudos (utilizados para apoiar a ABH) não descrevem um único padrão de atenção selectiva., Um outro ponto importante a considerar é a própria definição de familiaridade e associação de grupos adotada pela maioria dos estudos utilizados para apoiar a ABH (Massen e Gallup, 2017). Estes estudos mostraram uma melhor detecção visual e codificação visual perceptiva de rostos de sujeitos familiares/em grupo em comparação com os não familiares, mas definiram familiaridade e filiação em grupo não com base nas relações pessoais entre indivíduos. Em vez disso, a familiaridade ou a filiação em grupo foram definidas com base no conhecimento indireto (e.g.,, foto de pessoas famosas ou de um assunto já mostrado em fases pré-julgamento) ou raça comum (por exemplo, Buttle e Raymond, 2003; Ganel e Goshen-Gottstein, 2004; Jackson e Raymond, 2006; Michel et al., 2006). Esta definição é adequada para os propósitos destes estudos, mas não é, como muito bem se os resultados são utilizados para propor explicações alternativas para a influência que a verdadeira ligação social baseada em relações reais—pode ter sobre um fenômeno, neste caso, o contágio do bocejo. Por exemplo, Michel et al., (2006) observou-se que os indivíduos caucasianos e asiáticos podiam reconhecer melhor rostos de mesma raça, mas esta diferença não estava presente em indivíduos asiáticos que viviam há cerca de um ano entre caucasianos. Outro ponto de discussão diz respeito à presença de contágio do bocejo em crianças com transtorno do espectro do autismo (ASD), que freqüentemente mostram alterações na atenção visual (Richard e Lajiness-O’Neill, 2015). Nas crianças da ASD, o contágio bocejante pode estar ausente (Senju et al., 2007), imparidade (Helt et al.,, 2010), ou semelhante ao desenvolvimento típico de crianças quando os sujeitos são induzidos a redirecionar sua atenção ao estímulo de vídeo durante os ensaios experimentais (Usui et al., 2013). Num estudo recente, Mariscal et al. (2019) descobriu que o contágio de bocejos em crianças com DSA estava positivamente relacionado com a concentração de oxitocina no sangue, a hormona envolvida no apego parental e social (Decety et al., 2016) e postulou que o contágio bocejante em crianças ASD pode estar relacionado a concentrações médias variáveis de ocitocina em diferentes coortes de estudo (Mariscal et al., 2019)., Esta descoberta está em linha com a hipótese da EBH que liga taxas de contágio bocejadas à ligação social, que pode refletir ligação emocional.
Conclusion
Our study adds to the discussion over the mechanisms under the social asymmetry in yawn contagion (for a critical reviews: see Adriaense et al., 2020; Palagi et al., 2020), mostrando que o contágio bocejante está provavelmente associado com a bottom-up, em vez de com a atenção seletiva de cima para baixo., A atenção ascendente é principalmente liderada pela percepção sensorial do estímulo eliciante, enquanto a atenção seletiva de cima para baixo é um processo voluntário e sustentado no qual um determinado item é selecionado internamente e focado ou examinado (Katsuki e Constantinidis, 2014). A este respeito, o estímulo acústico (bocejo auditivo) emitido pelo gatilho foi ouvido e poderia provocar uma resposta bocejante no receptor, mesmo que o receptor não estivesse prestando qualquer atenção visual voluntária ao gatilho., Além disso, as taxas de resposta bocejante foram socialmente moduladas, com o contágio auditivo bocejando sendo mais alto em indivíduos que estavam mais fortemente ligados um ao outro. Assim, a atenção seletiva de cima para baixo não é o principal motor da assimetria social observada no contágio de bocejo, que parece ser um fenômeno impulsionado pelo estímulo relacionado com os processos de atenção de baixo para cima. É necessária uma investigação mais aprofundada para compreender se e de que forma outras formas de atenção ou estágios pré-atentos são capazes de afetar o contágio do bocejo.,o conjunto de dados utilizado para este estudo é fornecido no material suplementar.o estudo envolvendo participantes humanos foi revisado e aprovado pelo Comitato di Bioetica D’Ateneo (Comitê de Bioética da Universidade)—Universidade de Turim (ref. no. 451945). Para este estudo, não era necessário um consentimento informado e escrito para a participação, em conformidade com a legislação nacional e os requisitos institucionais.
contribuições dos autores
In E EP realizou a coleta de dados e concebeu e escreveu o manuscrito., AZ ajudou na coleta de dados e revisão de manuscritos, figuras e tabelas. MG realizou análises estatísticas e escreveu a parte relacionada do manuscrito.
conflito de interesses
os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.os autores desejam agradecer ao Prof. Roberto Barbuti e à Prof. Cristina Giacoma por terem apoiado a pesquisa na Universidade de Pisa e na Universidade de Turim, respectivamente.,
Supplementary Material
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–266. doi: 10.1163/156853974×00534
PubMed Abstract | CrossRef Full Text | Google Scholar
Anderson, J. R., and Meno, P. (2003). Psychological influences on yawning in children. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J., and Cirulli, E. T. (2014)., A variação Individual da susceptibilidade ao bocejo contagioso é altamente estável e largamente inexplicável por empatia ou outros factores conhecidos. PLoS One 9: e91773. doi: 10.1371 / journal.pone.0091773
PubMed Abstract | CrossRef Full Text/Google Scholar
Bretz, F., Hothorn, T., and Westfall, P. (2010). Multiple Comparisons Using R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, B. L. (1994). Estudo etológico do bocejo em primatas. I., Quantitative analysis and study of causation in two species of Old World monkeys (Cercocebus albigena and Macaca fascicularis). Ethology 98, 221-245. doi: 10.1111 / J. 1439-0310. 1994.tb01073.X
CrossRef Full Text | Google Scholar
Dobson, A. J. (2002). An Introduction to Generalized Linear Models, 2nd Edn. Boca Raton, FL: Chapman and Hall.
Google Scholar
Katsuki, F., and Constantinidis, C. (2014). Atenção de baixo para cima e de cima para baixo: diferentes processos e sistemas neurais sobrepostos., Neurocientista 20, 509-521. doi: 10.1177 / 1073858413514136
PubMed Abstract | CrossRef Full Text/Google Scholar
Singer, T. (2006). The neuronal basis and ontogeny of empathy and mind reading: review of literature and implications for future research. Neuroci. Biobehav. Rev. 30, 855-863. doi: 10.1016 / j. neubiorev.2006.06.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Statistics Canada (2009). Categorias Etárias, Grupos De Ciclo De Vida. Ottawa: Statistics Canada.,
Google Scholar