Frontiere în Psihologie
Introducere
Căscatul este un involuntare secvență de deschidere gura, inspirație profundă, scurtă apnee, și mai mult sau mai puțin lent de expirare (Baenninger, 1997; Walusinski și Deputte, 2004; Guggisberg et al., 2010; Krestel și colab., 2018). Când este provocat, un căscat nu poate fi complet suprimat. Prin urmare, a fost definit ca un model stereotip sau reflex (Lehmann, 1979; Provine, 1986)., În Homo sapiens, mai multe ipoteze au fost puse mai departe cu variabila de sprijin pentru a explica mecanismele și funcțiile spontan căscat, cum ar fi oxigenarea (funcția respiratorie cauzată de hipoxie), legate de stres comportament (cauzate de excitare), sau de termoreglare (cauzate de hipertermie; Guggisberg et al., 2010; Massen și colab., 2014; Gallup și Gallup, 2019). Fiind un răspuns fiziologic, căscatul poate fi afectat de factori interni și externi, cum ar fi timpul zilei (Giganti și Zilli, 2011) sau temperatura intracraniană/cerebrală (Gallup și Eldakar, 2013).,căscatul poate fi auto-direcționat și / sau afișat altora (Moyaho et al., 2017; Palagi și colab., 2019). La primatele umane și non-umane, în funcție de specie, atunci când căscatul este arătat altora, poate comunica amenințare (Troisi et al., 1990; deputat, 1994) și / sau schimbări fiziologice și comportamentale (Provine et al., 1987; Leone și colab., 2015; Zannella și colab., 2015). La om, căscatul este un răspuns modulat social, deoarece poate fi inhibat de prezența socială reală și nu virtuală (Gallup et al.,, 2019) și pentru că un căscat poate fi declanșat de căscatul altcuiva, ca urmare a unui fenomen cunoscut sub numele de căscat contagios (Provine, 1989, 2005). Contagiunea căscatului poate fi provocată chiar dacă căscatul este auzit, dar nu este văzut (Arnott et al., 2009; Massen și colab., 2015).
oamenii lor filogenetic mai apropiate ape specii (cimpanzei: Pan paniscus; bonobo: Pan troglodytes) și maimuță Africană Theropithecus gelada, contagios căscatul nu este numai prezent (Provine, 1986; Palagi et al., 2009; Tan și colab., 2017; dar vezi: Amici și colab.,, 2014), dar și modulată social, deoarece răspunsul căscat este cel mai mare atunci când sunt implicate anumite categorii de indivizi (de exemplu, rude, membri de grup, dominanți; Palagi et al., 2009; Campbell și de Waal, 2011, 2014; Norscia și Palagi, 2011; Demuru și Palagi, 2012; Massen et al., 2012)., Două argumente principale au fost prezentate pentru a explica această asimetrie socială în contagios căscatul, care au fost grupate în două ipoteze principale: Emoțională Părtinire Ipoteza (EBH), care leagă contagios căscatul emoționale transfer, și de distorsiune Atențională Ipoteza (ABH), care consideră că contagios căscatul ca pe un motor de răspuns care este supusă diferențe în atențional top-down procese (Palagi et al., 2020).,EBH prezice că asimetria socială observată în ratele de contagiune a căscatului reflectă diferențele în diferitele legături sociale, un proxy al legăturii emoționale, între indivizi. Această ipoteză este susținută de dovezi că ratele de contagiune a căscatului urmează o tendință empatică (sensu Preston și de Waal, 2002), fiind cea mai mare dintre indivizii care împărtășesc o legătură emoțională mai puternică. Mai exact, Norscia and Palagi (2011) a constatat că la om ratele de contagiune căscat sunt mai mari ca răspuns la rude și prieteni decât ca răspuns la cunoștințe și străini., În adult cimpanzei molipsirea căscatului este mai mare între in-group față de out-grup de membri (Campbell și de Waal, 2011) și în bonobo căscatul e contagios, ratele sunt mai mari între persoane afiliate mai mult unul cu altul (Demuru și Palagi, 2012). Într-o investigație comparativă care include atât oameni, cât și bonobos, Palagi și colab. (2014) a constatat că ratele de contagiune a căscatului au fost afectate de calitatea relației dintre indivizi mai mult decât de speciile de care aparțineau subiecții., În plus, la om, contagiunea căscării crește odată cu vârsta, când capacitatea de a identifica emoțiile altora crește și scade odată cu vârsta înaintată, când o astfel de abilitate scade (Wiggers și van Lieshout, 1985; Anderson și Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen și Anderson, 2011; Bartolomeu și Cirulli, 2014). Ratele de contagiune a căscatului cresc de la copilărie până la maturitate, de asemenea, la cimpanzei (Madsen și Persson, 2013).ABH prezice că asimetria socială observată în contagiunea căscatului se poate datora diferențelor de atenție socială, vizuală (Massen și Gallup, 2017)., În special, cele mai înalte niveluri de contagios căscatul ar fi din cauza extra de sus în jos, selectiv vizuale atenție la persoanele care sunt mai relevante pentru observator, cum ar fi subiecte familiare, ca apare la om și maimuțele, sau dominante, așa cum apare în cimpanzei și bonobo (Yoon și Vania, 2010; Massen et al., 2012; Massen și Gallup, 2017). Potrivit lui Massen and Gallup( 2017), ABH ar fi susținută de dovezile existente privind detectarea vizuală diferită și codificarea perceptivă vizuală a fețelor subiecților familiari și/sau din grup comparativ cu cei necunoscuți (de exemplu.,, Buttle și Raymond, 2003; Ganel și Goshen-Gottstein, 2004; Jackson și Raymond, 2006; Michel et al., 2006).în acest studiu, am analizat datele despre căscat colectate pe parcursul a 9 ani pe oameni în setările lor naturale și am extrapolat cazurile în care căscatul emis de un subiect ar putea fi auzit, dar nu văzut de un potențial răspuns (căscat auditiv). Luând în considerare numai cazurile în care semnalul vizual al stimulului căscat nu a fost detectabil, am verificat dacă asimetria socială observată anterior în ratele de contagiune a căscatului a persistat sau nu., În special, am testat următoarele predicții alternative derivate din cele două ipoteze prezentate mai sus (EBH și ABH).
Predicția 1a: conform EBH, ratele de contagiune a căscatului sunt influențate de puterea legăturii sociale interindividuale-un proxy al legăturii emoționale—în sine și nu de o atenție vizuală selectivă de sus în jos, acordată anumitor indivizi în special., Dacă această ipoteză este susținută, ne așteptăm să respecte prejudecată socială, de asemenea, atunci când tac vizual a căscat stimul este exclusă și ratele de auditiv contagios căscatul să fie mai mare între puternic legat față de slab legat persoane fizice.Predicția 1B: conform ABH, nivelurile mai ridicate de contagiune a căscatului între persoanele puternic legate în comparație cu persoanele slab legate ar fi legate de cea mai apropiată atenție vizuală selectivă de sus în jos pe care indivizii o plătesc persoanelor care sunt relevante pentru ei, de exemplu, familia și prietenii., Dacă această ipoteză este susținută, părtinirea socială observată în contagiunea căscatului ar trebui să dispară atunci când sunt luate în considerare doar căscăturile auditive, deoarece semnalul vizual nu poate fi urmat de potențialul respondent.
Materiale și Metode
Colectarea de Date și Definițiile Operaționale
Pentru acest studiu, am considerat vocalized cască emisă de un subiect care putea fi auzit doar—dar nu a văzut—de un potențial răspuns (denumită în continuare: auditive cască). Emițătorul și potențialii respondenți au trebuit să fie într-un interval de ≤5 m., Vocalized cască implicat utilizarea de falduri vocale și cască că numai implicați grele inspirație/expirare nu au fost considerate ca vocalized; 294 de cazuri de auditiv casca au fost extrapolate de la un set de date dintr-un total de 2001 căscat crize colectate peste 9 ani—din 2010 până în 2019—folosind toate aparițiile metodă de eșantionare (Altmann, 1974). Mai exact, căscăturile auditive au fost colectate din noiembrie 2010 până în Mai 2019, între orele 05.30 și 02.30, pe subiecți caucazieni umani, cu vârste cuprinse între 18 și 77 de ani, în timpul activităților lor de rutină, de exemplu, în locuri de muncă, peste mese, în timpul întâlnirilor sociale etc.,, subiecții fiind conștienți de a fi observați și în absența oricărei surse externe evidente de anxietate. Baza de date a căscatului auditiv a inclus 193 de diade de răspuns potențial de răspuns. În funcție de situație, informațiile au fost înregistrate, notate, prin coduri alfanumerice și introduse direct în foi de calcul, tastate în telefoane mobile sau scrise pe hârtie, apoi introduse în foi de calcul pentru elaborarea ulterioară. Informațiile de bază, cum ar fi vârsta și relația dintre oameni, au fost cunoscute autorilor., Potențialul de răspuns au fost codificate ca în non-vedere starea când capul lor a fost rotit cu 180°, cu privire la declanșare sau când o fizic, vedere blocarea obstacol a fost prezent prevenirea potențialul de răspuns de la a vedea pe trăgaci fața și corpul. Declanșatorul și respondentul nu au fost niciodată complet izolate (de exemplu, în două camere separate, cu uși închise) una de cealaltă., Social apropierea fost colectate de pe patru niveluri: 0 = străini, care nu au întâlnit înainte; 1 = cunoștințe, care împărtășită exclusiv o relație indirectă, bazată pe un al treilea element extern, care este de muncă (colegii) sau prieteni în comun (prieteni de prieteni); 2 = prietenii, non-legate de persoane împart o relație directă nu exclusiv legate de un al treilea element extern; 3 = regulat parteneri și rude (r ≥ 0.25)., Literatura anterioară raportează că răspunsurile la căscat pot fi provocate în 5 minute după ce au urmărit căscatul altcuiva (căscatul declanșatorului) (Provine, 1986), cu un maxim în primul minut (Provine, 2005; Palagi et al., 2014). Literatura de specialitate raportează, de asemenea, că la cel de-al patrulea minut există o mai mare probabilitate de autocorelație (în sensul că prezența unui căscat efectuate de către un subiect la t0 crește probabilitatea de a avea un alt căscat de același subiect la t(0+X), unde X este creșterea unitatea de timp; Kapitány și Nielsen, 2017)., Prin urmare, am considerat răspunsurile căscatului care apar într-o fereastră de timp de 3 minute de la căscatul emis de declanșator. Pentru a reduce și mai mult prejudecata autocorelației, în cazul unui lanț de căscări emise de declanșator (mai multe căscări emise în fereastra de timp 3-min) am înregistrat ca răspuns doar primul căscat efectuat după percepția ultimului căscat. Am codificat un căscat ca fiind „spontan” atunci când niciun alt subiect nu a căscat în cele 5 minute anterioare evenimentului căscat., codat ca: 1 = prezența, 0 = absenta; sociale bond a fost introdus cu patru niveluri definite mai sus (0 = străini; 1 = cunoștințe; 2 = prieteni; 3 = kin); de declanșare și observator de sex au fost etichetate ca fiind: M = masculin, F = feminin; vârsta clase de declanșare și de răspuns au fost codificate după cum urmează: yo = tineri (18-24 ani); ad = adulte (25-64 ani); se = seniori (peste 65 ani) (Statistici Canada, 2009); sloturi de timp au fost codificate după cum urmează: 1 = 05:30-09:00; 2 = 09:01 am–12:30 pm; 3 = 12:31-16:00 pm; 4 = 16:01-19:30 pm; 5 = 19:31-23:00 pm; 6 = 23:01-02:30 (Giganti si Zilli, 2011)., Baza de date (a se vedea fișa tehnică suplimentară) a inclus 84 de bărbați, 69 de femei, 16 tineri (yo), 122 de adulți (ad) și 15 seniori (se)., Pentru a testa dacă apariție a căscat contagios a fost influențată de factori de obligațiuni (0 = străini; 1 = cunoștințe; 2 = prieteni; 3 = kin), sex de declanșare (Trigger_sex), sexul răspuns (Responder_sex), vârsta, clasa de declanșare (Trigger_ageclass), iar clasa de varsta a răspuns (Responder_ageclass), și slot de timp (de la 1 la 6), am folosit un liniar generalizat model mixt (GLMM) care a inclus aceste cinci predictori ca efecte fixe și factori declanșatori (Trigger) si de interventie (Raspuns) identitatea ca efecte aleatoare. Am montat modelele în R (R Core Team, 2018; versiunea 3.5.,1) Folosind funcția Lmer a pachetului R lme4 (Bates și colab., 2015). Am stabilit semnificația modelului complet prin comparație cu un model nul care cuprinde doar efectele aleatorii (Forstmeier și Schielzeth, 2011). Am folosit un test de probabilitate (Dobson, 2002) pentru a testa această semnificație (ANOVA cu argumentul „Chisq”). Am calculat valorile p pentru predictorii individuali pe baza testelor raportului de probabilitate între modelul complet și modelul nul folosind funcția R „drop1” (Barr et al., 2013). Deoarece variabila de răspuns a fost binară, am folosit o distribuție de eroare binomială., Am testat dacă interacțiunea dintre sexe sau clasele de vârstă ale declanșatorului și respondentului au fost semnificative, dar, deoarece nu au fost, nu le-am inclus în model. Am folosit un pachet de contrast multiplu (multcomp) pentru a efectua toate comparațiile perechi pentru fiecare nivel de lipire cu testul Tukey (Bretz et al., 2010). Am raportat valorile P ajustate la Bonferroni, estimarea (Est), eroarea standard (SE) și valorile Z.
Rezultate

Figura 1., Linia de complot a efectului legăturii sociale dintre declanșator și responder (axa X) asupra apariției medii a contagiunii căscate acustice (axa Y). Prietenii și rudele arată semnificativ mai mare căscat contagios frecvențe decât străini și cunoștințe (testul Tukey: prieteni față de străini p < 0.001; kin față de străini p < 0.001; prietenii versus cunoștințe p < 0.001; kin versus cunoștințe p < 0.001; alte combinații, ns).,
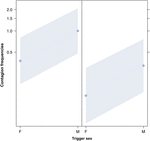
Figura 2. Linia de complot a efectului sexului declanșatorului (axa X) asupra incidenței medii a căscatului acustic (axa Y). Efectul sexului declanșator asupra contagiunii căscatului acustic atunci când respondentul este o femeie (dreapta) și un bărbat (stânga). Masculi, cască obtine mai mult cască decât femelele’ cele indiferent de sex a răspuns (rezultat al GLMM, p = 0.022). Punctele reprezintă efectul pe care variabila de răspuns îl are asupra variabilei independente „sexul declanșatorului”, pe baza valorii prezise de model., Benzile colorate arată intervalul de încredere de 95%.
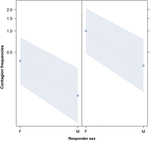
Figura 3. Linia de complot a efectului sexului respondentului (axa X) asupra incidenței medii a căscatului acustic (axa Y). Efectul sexului respondent asupra contagiunii căscatului acustic atunci când declanșatorul este o femeie (dreapta) sau un bărbat (stânga). Femelele răspund semnificativ mai mult decât bărbații, indiferent de sexul declanșatorului (rezultatul GLMM, p = 0,021)., Punctele reprezintă efectul pe care variabila de răspuns îl are asupra variabilei independente „sexul respondentului”, pe baza valorii prezise de model. Benzile colorate arată intervalul de încredere de 95%.acest studiu arată pentru prima dată că contagiunea căscatului este afectată în mod semnificativ de legătura socială dintre indivizi (Tabelul 1) chiar și atunci când stimulii declanșatori sunt căscări auditive, pe care le-am definit ca căscări vocalizate care ar putea fi auzite, dar nu văzute (cue vizual nedetectabil, cue auditiv detectabil)., În special, căscatul contagios auditiv este semnificativ mai frecvent între rude și prieteni decât între străini și cunoscuți (Figura 1). Această constatare susține predicție 1a bazează pe EBH și nu de predicție 1b bazează pe ABH, ceea ce duce la concluzia că la oameni de sus în jos, selectiv atenția vizuală nu poate fi principalul factor de asimetrie socială observată în căscatul e contagios rate (Norscia și Palagi, 2011; Norscia et al., 2016a)., De asemenea, sexul declanșatorului și sexul receptorului au avut un efect semnificativ asupra ratelor de contagiune a căscatului, bărbații—ca declanșatori—răspunzând de alții mai frecvent decât femeile (Figura 2), iar femeile răspunzând mai frecvent la căscatul altora decât bărbații (Figura 3).spre deosebire de Bartolomeu și Cirulli (2014), nu am găsit niciun efect de vârstă asupra contagiunii căscatului, cel mai probabil pentru că baza noastră de date privind căscatul auditiv a avut o prevalență puternică a adulților (25-64 de ani)., Cele mai înalte niveluri de contagiune a căscatului auditiv la femei în comparație cu bărbații confirmă părtinirea de gen observată în condiții naturaliste asupra oamenilor susceptibili la contagiunea căscatului de către Norscia et al. (2016a, b) atunci când se analizează un set de date mai mare, care a inclus, de asemenea, căscatul care ar putea fi văzut de respondentul potențial (cu modalitatea senzorială căscat—viziune, auz sau ambele—care nu afectează răspunsul)., Prejudecata de gen este, de asemenea, în acord parțial cu rezultatele anterioare obținute în setări controlate, inclusiv cue vizual (Chan și Tseng, 2017; dar vezi Norscia și Palagi, 2011; Bartolomeu și Cirulli, 2014). S-a emis ipoteza că gradul ridicat de contagiune a căscatului la femei ar putea informa contagiunea emoțională (Norscia et al., 2016a), având în vedere abilitățile empatice relatărilor mai mari—legate de maternitate—ale femeilor în comparație cu bărbații (Christov-Moore et al., 2014)., Cu toate acestea, această problemă este încă în dezbatere, deoarece diferențele culturale dintre societățile umane pot modela dinamica legăturilor sociale într-un mod diferit. Prin urmare, este complicat, în această etapă a cunoașterii, să se descopere factorii culturali, calitatea relațiilor inter-personale și influența de gen în distribuția contagiunii căscatului. Rezultatele noastre arată, de asemenea, că bărbații au performanțe mai bune decât femeile ca declanșatori, iar ipoteza cea mai parsimonioasă pentru acest lucru ar putea fi că vocalizările bărbaților pot fi auzite mai bine în setări naturale, adesea caracterizate prin zgomote de fundal., Într-adevăr, percepția voce de gen se bazează în primul rând pe frecvența fundamentală, care este, în medie, mai mici cu o octavă în sex masculin decât de sex feminin voci, cu o frecvență mai mică vocalize calatori mai departe de înaltă frecvență (de Jder și Marler, 1977; Latinus și Taylor, 2012). Cu toate acestea, din cunoștințele noastre, nu există un studiu specific care să abordeze posibila părtinire de gen în audibilitatea căscatului, iar investigarea ulterioară cu studii experimentale în stare controlată este, prin urmare, necesară pentru a verifica această speculație.,în acest studiu, am constatat, de asemenea, că diferențele dintre ratele de contagiune a căscatului pe categorii (familie și prieteni, străini și cunoștințe) nu pot fi explicate prin diferențe în atenția vizuală selectivă de sus în jos. Această constatare este în conformitate cu literatura anterioară. Căscatul contagios pare să implice zone ale creierului care sunt mai mult legate de rețeaua de orientare de jos în sus decât de sus în jos . Mai mult, contagiunea căscatului nu este nici sensibilă la indiciile senzoriale prezente în semnal (auditiv, vizual sau audio-vizual) (Arnott et al.,, 2009; Norscia și Palagi, 2011), nici afectat de vedere vizual de declanșarea stimulului (cască în orientările de 90°, 180° și 270° sunt capabili să declanșeze căscat răspunsuri ca frontală, 0° cască; Provine, 1989, 1996). Chan and Tseng (2017) a constatat că abilitatea de a detecta un căscat ca atare (sensibilitatea la detectarea perceptuală) a fost legată de durata privirii la ochii stimulului care eliberează fața, dar modelele de ochi nu au putut modula căscatul contagios., La cimpanzei, frecvențele contagioase de căscat au fost cele mai ridicate între același grup decât indivizii din grupuri diferite, chiar dacă respondenții s-au uitat mai mult la videoclipurile cimpanzeilor din grup (Campbell și de Waal, 2011). Argumentul potrivit căruia atenția selectivă vizuală poate influența ratele de contagiune a căscatului într-o direcție specifică (subiecții care răspund mai mult familiei decât străinii) este, de asemenea, subminat de absența oricărui model specific de atenție socială în primatul uman și non-uman. Prin urmărirea ochilor (aplicată pentru a măsura timpul de vizualizare) și prin afișarea unor fețe necunoscute subiecților lor experimentali, Méary et al., (2014) a observat că oamenii au fost înclinați spre fețele de rasă proprie, în timp ce atenția macacilor rhesus a fost atrasă mai mult de fețele noi decât de aceleași specii. Kawakami și colab. (2014) a observat că subiecții umani au acordat mai multă atenție ochilor membrilor grupului etnic și nasului și gurii membrilor grupului etnic. Același studiu a arătat, de asemenea, că atenția vizuală nu depinde de rasa țintă. Măsurând cât timp subiecții experimentali au privit ecranul, Whitehouse et al., (2016) a observat că macacii Berberi mai atent la zgarieturi videoclipuri non-străin străin decât persoanele fizice, ci, de asemenea, remarcat faptul că în cadrul non-străini, macaci acordă mai multă atenție la persoanele cu care au împărțit o slabă relaționare socială. Prin măsurarea vedere ratele, Schino și Sciarretta (2016) a observat că mandrills uitat mai mult la propriile lor rude decât la non-rude, dar, de asemenea, mai dominant decât la subordonat grupului de prieteni. Prin urmare, aceste studii (utilizate pentru a sprijini ABH) nu descriu un singur model de atenție selectivă., Un alt punct important de luat în considerare este însăși definiția familiarității și apartenenței la grup adoptată de majoritatea studiilor utilizate pentru a sprijini ABH (Massen și Gallup, 2017). Aceste studii au arătat o mai bună detectare vizuală și o codificare perceptivă vizuală a fețelor subiecților familiari/în grup comparativ cu cei necunoscuți, dar au definit familiaritatea și apartenența la grup nu pe baza relațiilor personale dintre indivizi. În schimb, familiaritatea sau apartenența la grup au fost definite pe baza cunoștințelor indirecte (de exemplu,,, fotografii de oameni celebri sau de un subiect arătat deja în pre-proces de faze) sau rasa comună (de exemplu, Buttle și Raymond, 2003; Ganel și Goshen-Gottstein, 2004; Jackson și Raymond, 2006; Michel et al., 2006). Această definiție este bine pentru scopuri de aceste studii, dar nu este la fel de mult bine dacă rezultatele sunt utilizate pentru a propune explicații alternative pentru influența pe care real relații sociale bazate pe relații reale—pot avea asupra unui fenomen, în acest caz, căscatul e contagios. De exemplu, Michel și colab., (2006) a observat că subiecții caucazieni și asiatici ar putea recunoaște mai bine fețele de aceeași rasă, dar această diferență nu a fost prezentă la subiecții asiatici care trăiau de aproximativ un an printre caucazieni. Un alt punct de discuție se referă la prezența contagiunii căscatului la copiii cu tulburare de spectru autist (ASD), care prezintă frecvent modificări ale atenției vizuale (Richard și Lajiness-O ‘ Neill, 2015). La copiii cu TSA, contagiunea căscatului poate fi absentă (Senju et al., 2007), depreciate (Helt și colab.,, 2010), sau similar cu copiii care se dezvoltă în mod obișnuit atunci când subiecții sunt induși să-și redirecționeze atenția asupra stimulului video în timpul studiilor experimentale (Usui et al., 2013). Într-un studiu recent, Mariscal și colab. (2019) a constatat că contagiunea căscatului la copiii cu TSA a fost legată pozitiv de concentrația sanguină a oxitocinei, hormonul implicat în atașamentul parental și social (Decety et al., 2016) și a afirmat că contagiunea căscatului la copiii cu TSA poate fi legată de concentrațiile medii variabile de oxitocină în diferite cohorte de studiu (Mariscal et al., 2019)., Această constatare este în concordanță cu ipoteza EBH care leagă ratele de contagiune a căscatului de legătura socială, care poate reflecta legătura emoțională.
concluzie
studiul nostru adaugă la discuția asupra mecanismelor care stau la baza asimetriei sociale în contagiunea căscatului (pentru recenzii critice: vezi Adriaense et al., 2020; Palagi și colab., 2020), arătând că contagiunea căscatului este probabil asociată cu o atenție selectivă de jos în sus, mai degrabă decât cu o atenție selectivă de sus în jos., De jos în sus, atenția se duce în primul rând de percepția senzorială a provoca un stimul întrucât sus-jos, atenția selectivă este voluntară, susținută proces în care un anumit element este selectat pe plan intern și s-a axat pe sau examinate (Katsuki și Constantinidis, 2014). În acest sens, stimulul acustic (căscatul auditiv) emis de declanșator a fost auzit și ar putea provoca un răspuns căscat în receptor, chiar dacă receptorul nu acorda nicio atenție vizuală voluntară declanșatorului., Mai mult, ratele de răspuns la căscat au fost modulate social, contagiunea căscată auditivă fiind cea mai mare la persoanele care au fost cel mai puternic legate între ele. Prin urmare, atenția selectivă de sus în jos nu este principalul motor al asimetriei sociale observate în contagiunea căscatului, care pare a fi un fenomen stimulat-legat de procesele de atenție de jos în sus. Este necesară o investigație suplimentară pentru a înțelege dacă și în ce Mod alte forme de atenție sau etape pre-atente sunt capabile să afecteze contagiunea căscatului.,
Declarația de disponibilitate a datelor
setul de date utilizat pentru acest studiu este furnizat în materialul suplimentar.
Etică Declarație
studiu care implică participanți umani au fost revizuit și aprobat de către Comitato di Bioetica d’Ateneo (Universitatea de Bioetică, Comitetul)—Universitatea din Torino (ref. 451945). Pentru acest studiu nu a fost necesar consimțământul informat în scris pentru participare, în conformitate cu legislația națională și cerințele instituționale.
contribuțiile autorului
în și pe Efectuat colectarea de date și conceput și a scris manuscrisul., AZ a ajutat la colectarea datelor și revizuirea manuscriselor, figuri și tabele. MG a efectuat analize statistice și a scris partea aferentă a manuscrisului.
Conflict de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Aprecieri
autorii doresc să-i mulțumesc Prof. dr. Roberto Barbuti și Prof. Cristina Giacoma pentru a fi sprijinit de cercetare de la Universitatea din Pisa și la Universitatea din Torino, respectiv.,
Supplementary Material
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–266. doi: 10.1163/156853974×00534
PubMed Abstract | CrossRef Full Text | Google Scholar
Anderson, J. R., and Meno, P. (2003). Psychological influences on yawning in children. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J., and Cirulli, E. T. (2014)., Variația individuală în susceptibilitatea contagioasă a căscatului este foarte stabilă și în mare parte inexplicabilă de empatie sau de alți factori cunoscuți. PLoS Unul 9: e91773. doi: 10.1371 / jurnal.pone.0091773
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Bretz, F., Hothorn, T., și Westfall, P. (2010). Comparații Multiple folosind R. Boca Raton, FL: Chapman & Hall.Google Scholar
deputat, B. L. (1994). Studiul etologic al căscării la primate. Eu., Analiza cantitativă și studiul cauzalității la două specii de maimuțe din Lumea Veche (Cercocebus albigena și Macaca fascicularis). Etologie 98, 221-245. doi: 10.1111 / j.1439-0310.1994.tb01073.x
CrossRef Textul Complet | Google Scholar
Dobson, A. J. (2002). O introducere în modele liniare generalizate, Edn 2nd. Boca Raton, FL: Chapman și Hall.
Google Scholar
Katsuki, F., și Constantinidis, C. (2014). Atenție de jos în sus și de sus în jos: diferite procese și sisteme neuronale suprapuse., Neurologul 20, 509-521. doi: 10.1177/1073858413514136
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Cantareata, T. (2006). Baza neuronală și ontogenia empatiei și citirii minții: revizuirea literaturii și implicațiile pentru cercetările viitoare. Neurosci. Biobehav. Rev. 30, 855-863. doi: 10.1016/j.neubiorev.2006.06.011
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Statistics Canada (2009). Categorii De Vârstă, Grupe De Ciclu De Viață. Ottawa: Statistici Canada.,
Google Scholar