enkel Anatomi av näthinnan av Helga Kolb
Helga Kolb
1. Översikt.
När en ögonläkare använder ett oftalmoskop för att titta in i ögat ser han följande syn på näthinnan (Fig. 1).

i mitten av näthinnan är synnerven, ett cirkulärt till ovalt vitt område som mäter ca 2 x 1,5 mm över. Från mitten av den optiska nerven utstrålar de stora blodkärlen i näthinnan. Cirka 17 grader (4.,5-5 mm), eller två och en halv skiva diametrar till vänster om skivan, kan ses den något ovalformade, blodkärlsfri rödaktig fläck, fovea, som ligger i mitten av området som kallas makula av ögonläkare.
Fig. 1. Näthinnan som kan ses genom en opthalmoscope
KLICKA HÄR för att se en animation (från iris till näthinnan) (Quicktime-film)
Ett cirkulärt område på ca 6 mm runt fovea anses vara den centrala näthinnan medan utöver detta är perifera näthinnan som sträcker sig ora serrata, 21 mm från centrum av näthinnan (fovea)., Den totala näthinnan är en cirkulär skiva av mellan 30 och 40 mm i diameter (Polyak, 1941; Van Buren, 1963; Kolb, 1991).
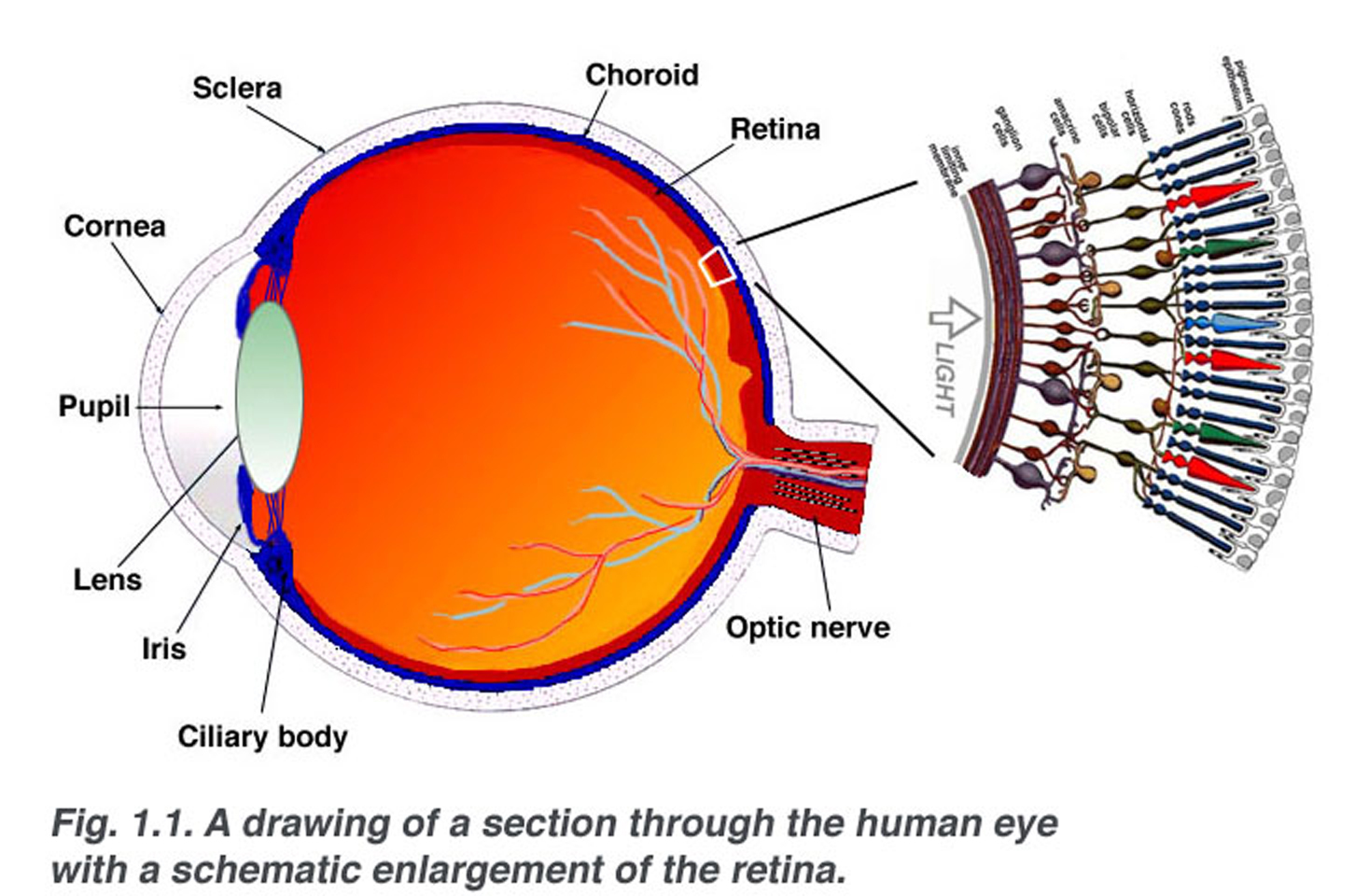
Fig. 1.1. En schematisk sektion genom det mänskliga ögat med en schematisk utvidgning av näthinnan
näthinnan är ungefär 0,5 mm tjock och linjer baksidan av ögat. Den optiska nerven innehåller ganglion cell axoner som löper till hjärnan och dessutom inkommande blodkärl som öppnar in i näthinnan för att vaskularisera retinala skikt och neuroner (Fig. 1.1)., En radiell sektion av en del av näthinnan avslöjar att ganglioncellerna (näthinnans utgående neuroner) ligger innerst i näthinnan närmast linsen och framsidan av ögat, och fotosensorerna (stavarna och konerna) ligger yttersta i näthinnan mot pigmentepitelet och choroid. Ljus måste därför resa genom näthinnans tjocklek innan du slår och aktiverar stavarna och konerna (Fig. 1.1)., Därefter översätts absorptionen av fotoner av fotoreceptorns visuella pigment till ett biokemiskt meddelande och sedan ett elektriskt meddelande som kan stimulera alla efterföljande neuroner i näthinnan. Det retinala meddelandet om den fototiska ingången och någon preliminär organisation av den visuella bilden i flera former av känsla överförs till hjärnan från ganglioncellernas spikmönster.,
ett förenklat kopplingsschema över näthinnan betonar endast de sensoriska fotoreceptorerna och ganglioncellerna med några interneuroner som förbinder de två celltyperna, såsom ses i Figur 2.
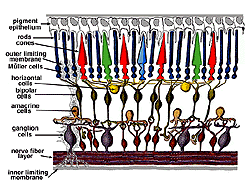
Fig. 2., Enkel organisation av näthinnan
när en anatomist tar en vertikal del av näthinnan och bearbetar den för mikroskopisk undersökning blir det uppenbart att näthinnan är mycket mer komplex och innehåller många fler nervcellstyper än det förenklade systemet (ovan) hade angivit. Det är omedelbart uppenbart att det finns många interneuroner förpackade i den centrala delen av näthinnan som ingriper mellan fotoreceptorerna och ganglioncellerna (Fig 3).,

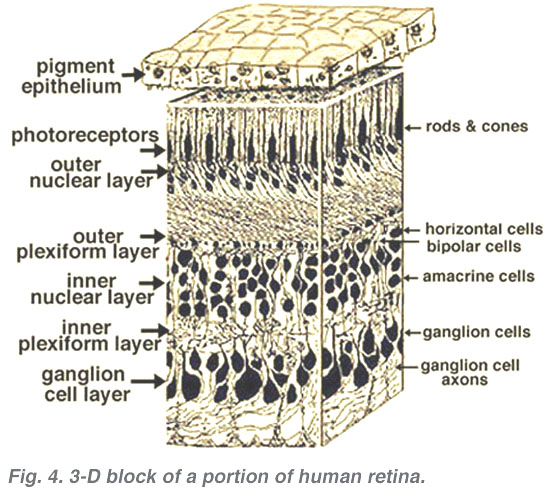
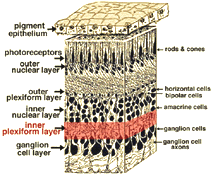
alla ryggradsdjur retinas består av tre lager av nervcellskroppar och två lager synapser (Fig. 4). Det yttre kärnskiktet innehåller cellkroppar av stavarna och konerna, det inre kärnskiktet innehåller cellkroppar av de bipolära, horisontella och amacrine-cellerna och ganglioncellskiktet innehåller cellkroppar av ganglionceller och förskjutna amacrine-celler. Dela dessa nervcellsskikt är två neuropils där synaptiska kontakter uppstår (Fig. 4).,
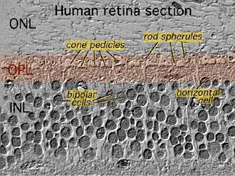
det första området av neuropil är det yttre plexiforma skiktet (OPL) där kopplingar mellan stång och kottar, och vertikalt kör bipolära celler och horisontellt orienterade horisontella celler uppträder (fikon. 5 och 6).
|
|
|

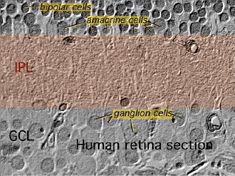
den andra neuropillen i näthinnan är det inre plexiforma skiktet (IPL), och det fungerar som en relästation för de vertikala informationsbärande nervcellerna, de bipolära cellerna, för att ansluta till ganglionceller (fikon. 7 och 8). Dessutom, olika sorter av horisontellt – och vertikalt riktade amacrine celler, på något sätt interagera i ytterligare nätverk för att påverka och integrera ganglion cellsignaler., Det är vid kulminationen av all denna neurala bearbetning i det inre plexiforma skiktet att meddelandet om den visuella bilden överförs till hjärnan längs den optiska nerven.
|
|
|
2. Centrala och perifera näthinnan jämfört.,
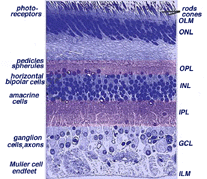
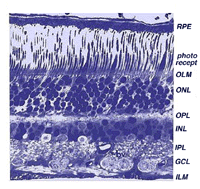
centrala näthinnan nära fovea är betydligt tjockare än perifera näthinnan (jämför fikon. 9 och 10). Detta beror på den ökade packningstätheten hos fotoreceptorer, särskilt konerna, och deras associerade bipolära och ganglionceller i centrala näthinnan jämfört med perifera näthinnan.
|
|
|
- Central näthinnan är kondominerad näthinna medan perifer näthinna är stångdominerad. Således i centrala näthinnan konerna är tätt åtskilda och stavarna färre i antal mellan konerna (fikon. 9 och 10).
- det yttre kärnskiktet (ONL), som består av cellkropparna i stavarna och konerna är ungefär samma tjocklek i centrala och perifera näthinnan., Men i det perifera stången cellkropparna överträffar kon cellkropparna medan det omvända är sant för centrala näthinnan. I centrala näthinnan har konerna sneda axoner som förskjuter sina cellkroppar från sina synaptiska pediklar i det yttre plexiforma skiktet (OPL). Dessa sneda axoner med åtföljande Muller cellprocesser bildar ett blekt färgande fibrösa utseende område som kallas Henle fiberskiktet. Det senare skiktet är frånvarande i perifer näthinna.,
- det inre kärnskiktet (INL) är tjockare i näthinnans centrala område jämfört med perifer näthinna, på grund av en större densitet av konanslutande andra ordningens neuroner (konbipolära celler) och mindre fält och närmare placerade horisontella celler och amacrine celler som berörs av konbanorna (Fig. 9). Som vi kommer att se senare är konanslutna kretsar av neuroner mindre konvergerande genom att färre koner impinge på andra ordningens neuroner, än stavar gör i stav-anslutna vägar.,
- en anmärkningsvärd skillnad mellan centrala och perifera näthinnan kan ses i de relativa tjocklekar av de inre plexiforma skikten (IPL), ganglion cellskikt (GCL) och nervfiberskikt (NFL) (fikon. 9 och 10). Detta beror återigen på det större antalet och den ökade packningstätheten hos ganglionceller som behövs för konbanorna i den kon-dominerande foveala näthinnan jämfört med den stångdominanta perifera näthinnan., Det större antalet ganglion celler innebär mer synaptisk interaktion i en tjockare IPL och större antal ganglion cell axoner coursing till synnerven i nervfiberskiktet (Fig. 9).
3. Muller gliaceller.
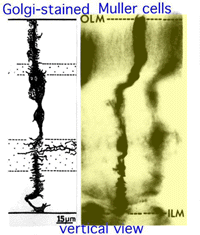
Fig. 11. Vertikal vy av Golgi färgade Muller glialceller
Muller celler är de radiella glialceller i näthinnan (Fig. 11). Det yttre begränsande membranet (OLM) i näthinnan bildas från vidhäftande korsningar mellan Mullerceller och fotoreceptorcellinnersegment., Det inre begränsande membranet (ILM) av näthinnan är jämväl sammansatt av lateralt kontakt Muller cell ändfötter och tillhörande basalmembrankomponenter.
OLM bildar en barriär mellan det subretinala utrymmet, i vilket de inre och yttre segmenten av fotoreceptorerna projektet att vara i nära samarbete med pigmentepitelskiktet bakom näthinnan, och den neurala näthinnan korrekt. ILM är den inre ytan av näthinnan som gränsar till glaskroppen och därigenom bildar en diffusionsbarriär mellan neural näthinnan och glaskroppen (Fig. 11).,
genom näthinnan levererar de stora blodkärlen i retinalkärlen de kapillärer som går in i nervvävnaden. Kapillärer finns som löper genom alla delar av näthinnan från nervfiberskiktet till det yttre plexiforma skiktet och även ibland så högt som i det yttre kärnskiktet. Näringsämnen från vaskulaturen av choriocapillaris (cc) bakom pigmentepitelskiktet levererar det känsliga fotoreceptorskiktet.
4. Foveal struktur.,
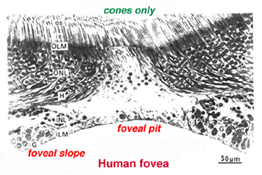
foveas centrum är känt som foveal grop (Polyak, 1941) och är en högspecialiserad region av näthinnan som skiljer sig från centrala och perifera näthinnan som vi hittills har övervägt. Radiella sektioner av denna lilla cirkulära region av näthinnan som mäter mindre än en fjärdedel av en millimeter (200 mikron) över visas nedan för människa (Fig. 12a) och för apa (Fig.12b).
|
|
|

fovea ligger i mitten av makulaområdet av näthinnan till den temporala sidan av det optiska nervhuvudet (Fig. 13a, A, B)., Det är ett område där konfotoreceptorer är koncentrerade vid maximal densitet, med undantag av stavarna, och anordnade vid sin mest effektiva packningstäthet som ligger i en sexkantig mosaik. Detta syns tydligare i en tangentiell sektion genom foveal kon inre segment (Fig. 13b).
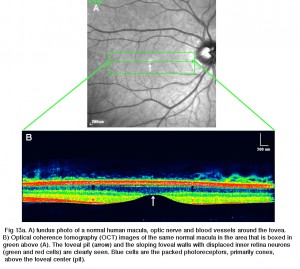
Fig 13a. a) fundus foto av en normal mänsklig makula, optisk nerv och blodkärl runt fovea. B) optisk koherens tomografi (OCT) bilder av samma normala makula i området som är inramat i grönt ovanför (a)., Foveal gropen (pilen) och de sluttande fovealväggarna med skingrade inre näthinnans neuroner (gröna och röda celler) ses tydligt. Blå celler är de packade fotoreceptorerna, främst kottar, ovanför foveal center (grop).

Fig. 13. Tangentiell sektion genom den mänskliga fovea
under denna centrala 200 mikron diameter centrala foveal grop, de andra skikten av näthinnan förskjuts koncentriskt lämnar endast det tunnaste arket av näthinnan bestående av koncellerna och några av deras cellkroppar (höger och vänster sida av fikon., 12a och 12b). Detta är särskilt väl ses i optisk koherens tomografi (OCT) bilder av levande öga och näthinna (Fig. 13a, B). Radiellt förvrängd men fullständig skiktning av näthinnan visas sedan gradvis längs foveal sluttningen tills kanten av fovea består av de förskjutna andra och tredje ordningens neuroner relaterade till de centrala konerna. Här staplas ganglioncellerna i sex lager så att detta område kallas fovealfälgen eller parafovea (Polyak, 1941), den tjockaste delen av hela näthinnan.
5. Macula lutea.,
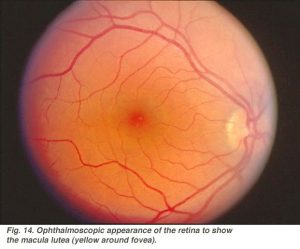
hela fovealområdet inklusive foveal pit, foveal slope, parafovea och perifovea anses vara makula i det mänskliga ögat. Bekant för ögonläkare är en gul pigmentering till makulaområdet som kallas macula lutea (Fig. 14).
 denna pigmentering är reflektionen från gula screeningpigment, xantofyllkarotenoiderna zeaxantin och lutein (Balashov och Bernstein, 1998), närvarande i Konaxonerna i Henlefiberskiktet. Macula lutea anses fungera som ett kort våglängdsfilter, utöver det som tillhandahålls av linsen (Rodieck, 1973)., Eftersom fovea är den viktigaste delen av näthinnan för mänsklig syn är skyddsmekanismer för att undvika starkt ljus och särskilt ultraviolett bestrålningsskada väsentliga. För, om de känsliga konerna i vår fovea förstörs blir vi blinda.
denna pigmentering är reflektionen från gula screeningpigment, xantofyllkarotenoiderna zeaxantin och lutein (Balashov och Bernstein, 1998), närvarande i Konaxonerna i Henlefiberskiktet. Macula lutea anses fungera som ett kort våglängdsfilter, utöver det som tillhandahålls av linsen (Rodieck, 1973)., Eftersom fovea är den viktigaste delen av näthinnan för mänsklig syn är skyddsmekanismer för att undvika starkt ljus och särskilt ultraviolett bestrålningsskada väsentliga. För, om de känsliga konerna i vår fovea förstörs blir vi blinda.
Fig. 14. Oftalmoskopisk utseende av näthinnan för att visa makula lutea

Fig. 15. Vertikal sektion genom apan fovea för att visa fördelningen av macula lutea. Från Snodderly et al.,, 1984
det gula pigmentet som bildar makula lutea i fovea kan tydligt demonstreras genom att visa en del av fovea i mikroskopet med blått ljus (Fig. 15). Det mörka mönstret i foveal gropen som sträcker sig ut till kanten av foveal-lutningen orsakas av den makulära pigmentfördelningen (Snodderly et al., 1984).

Fig. 16., Utseende av konmosaiken i fovea med och utan makula lutea
om man skulle visualisera foveal fotoreceptormosaiken som om de visuella pigmenten i de enskilda konerna inte blektades, skulle man se bilden som visas i figur 16 (nedre ram) (bild från Lall och Cone, 1996). De korta våglängdskänsliga konerna på foveal sluttningen ser blekgul grön ut, de mellersta våglängdskottarna, rosa och de långa våglängdskänsliga konerna, lila., Om vi nu lägger till effekten av det gula screeningpigmentet i makula lutea ser vi utseendet på konmosaiken i figur 16 (övre ram). Macula lutea hjälper till att förbättra akromatisk upplösning av fovealkottarna och blockerar skadlig UV-ljusbestrålning (Fig. 16 från Abner Lall och Richard Kon, opublicerade data).
6. Ganglion cell fiber lager.
ganglion cell axoner körs i nervfiberskiktet ovanför det inre begränsande membranet mot synnerven huvudet i en bågform (Fig. 00, strömmande rosa fibrer)., Fovea är naturligtvis fri från ett nervfiberskikt eftersom de inre näthinnan och ganglioncellerna skjuts bort till foveal sluttningen. De centrala ganglion-cellfibrerna löper runt foveal-lutningen och sveper i riktning mot den optiska nerven. Perifer ganglion cell axoner fortsätta denna båggång till synnerven med en dorso / ventral split längs den horisontella meridianen (Fig. 00). Retinal topografi upprätthålls i optisk nerv, genom det laterala geniculatet till den visuella cortexen.
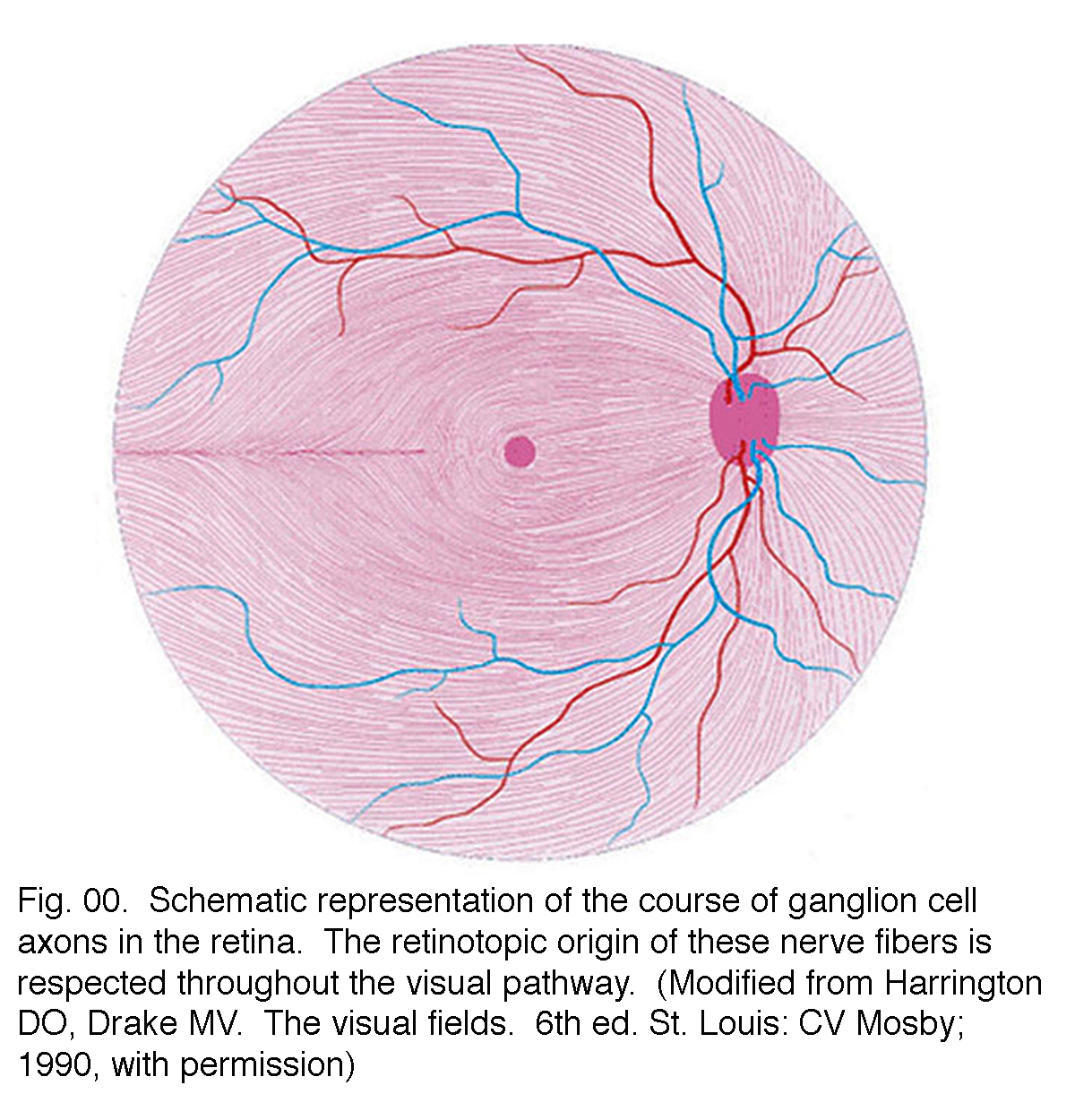
Fig. 00., Schematisk representation av ganglion cell axoner i näthinnan. Det retinotopiska ursprunget för dessa nervfibrer respekteras under hela den visuella vägen. (Modifierad från Harrington GÖRA, Drake MV. Synfält. 6th ed. St Louis: CV Mosby’; 1990, med tillstånd)
7. Blodtillförsel till näthinnan.
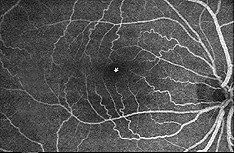
det finns två källor till blodtillförsel till däggdjursretina: den centrala retinalartären och de koroida blodkärlen. Choroid får det största blodflödet (65-85%) (Henkind et al.,, 1979) och är avgörande för upprätthållandet av den yttre näthinnan (särskilt fotoreceptorerna) och de återstående 20-30% strömmar till näthinnan genom den centrala retinala artären från det optiska nervhuvudet för att närma de inre retinala skikten. Den centrala retinalartären har 4 huvudgrenar i den mänskliga näthinnan (Fig. 17).
Fig. 17. Fundus fotografi visar flourescein avbildning av de stora artärer och vener i en normal mänsklig högra ögonhinnan., Fartygen kommer ut ur synnerven huvudet och kör på radiellt sätt böjda mot och runt fovea (asterisk i fotografi) (bild artighet av Isabel Pinilla, Spanien)
de arteriella intraretinala grenarna levererar sedan tre lager av kapillärnät dvs 1) de radiella peripapillära kapillärerna (RPCs) och 2) ett inre och 3) ett yttre lager av kapillärer (Fig. 18a). Prekapillära venoler dräneras i venoler och genom motsvarande vensystem till den centrala retinala venen (Fig. 18b).,
|
|
|
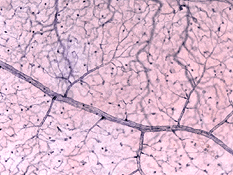
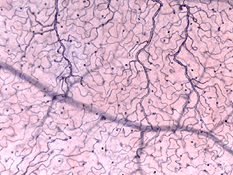
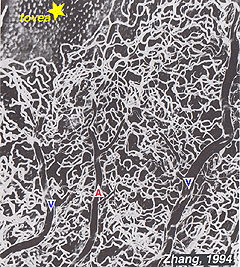
de radiella peripapillära kapillärerna (RPCs) är det mest ytliga skiktet av kapillärer som ligger i den inre delen av nervfiberskiktet och löper längs vägarna i de stora superotemporala och inferotemporala kärlen 4-5 mm från den optiska skivan (Zhang, 1994). RPCs anatomose med varandra och de djupare kapillärerna. De inre kapillärerna ligger i ganglion-cellskikten under och parallellt med RPCs., Det yttre kapillärnätet löper från det inre plexiforma skiktet till det yttre plexiforma skiktet trodde det inre kärnskiktet (Zhang, 1974).
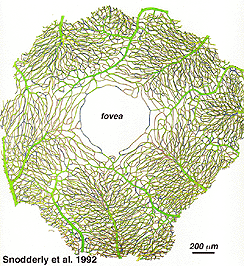
som kommer att märkas från flouresceinangiografi av figur 17, där som en ring av blodkärl i makulaområdet runt ett blodkärl-och kapillärfri zon 450-600 um i diameter, betecknar fovea. De makulära kärlen härrör från grenar av de överlägsna temporala och inferotemporala artärerna. Vid gränsen till den avaskulära zonen blir kapillärerna två skiktade och slutligen gå med som en enda skiktad ring., Uppsamlingsvenulerna är mer djupa (bakre) till arteriolerna och dränerar blodflödet tillbaka i huvudåren (Fig. 19, från Zhang, 1974). I rhesusapan syns denna perimakulära ring och blodkärlsfria fovea tydligt i de vackra ritningarna som gjorts av Max Snodderlys grupp (Fig. 20, Sodderly et al., 1992.)
|
|
|
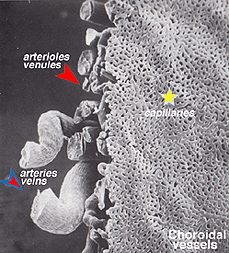
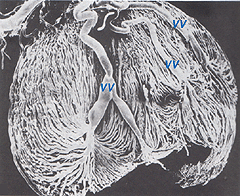
de koroidala artärerna uppstår från långa och korta bakre ciliära artärer och grenar av Zinns cirkel (runt den optiska skivan)., Var och en av de bakre ciliära artärerna bryts upp i fläktformade lobuler av kapillärer som levererar lokaliserade regioner av choroid (Hayreh, 1975). Makulaområdet hos de koroida kärlen är inte specialiserade som retinalblodförsörjningen är (Zhang, 1994). Artärerna genomborrar sklera runt synnerven och fläkt ut för att bilda de tre vaskulära skikten i åderhinnan: yttre (mest skleral), mediala och inre (närmaste Bruchs membran av pigmentepitelet) skikt av blodkärl. Detta visas tydligt i korrosionsgjutningen av ett snitt av den mänskliga choroiden i Figur 21a (Zhang, 1974)., De motsvarande venösa lobulerna dräneras i venulerna och venerna som löper fram mot ögonbollens ekvator för att komma in i virvelvenerna (Fig. 21b). En eller två vortex vener dränerar var och en av ögonbollens 4 kvadranter. Virvelvenerna tränger in i sclera och slår samman i den oftalmiska venen som visas i korrosionsgjutningen i Figur 21b (Zhang. 1994).
|
|
|
8., Degenerativa sjukdomar i den mänskliga näthinnan.
den mänskliga näthinnan är en känslig organisation av neuroner, glia och närande blodkärl. I vissa ögonsjukdomar blir näthinnan skadad eller komprometterad, och degenerativa förändringar som i så fall leder till allvarliga skador på nervcellerna som bär de vitala mesagerna om den visuella bilden till hjärnan. Vi indikerar fyra olika tillstånd där näthinnan är sjuk och blindhet kan vara slutresultatet. Mycket mer information om patologi i hela ögat och näthinnan finns på en webbplats gjord av ögonpatologen Dr., Nick Mamalis, Moran Eye Center.
|
|
|
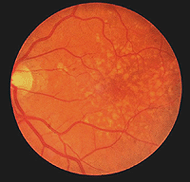
åldersrelaterad makuladegeneration är ett vanligt retinalproblem i åldrandet och en ledande orsak till blindhet i världen. Makulaområdet och fovea blir komprometterade på grund av pigmentepitelet bakom näthinnan som degenererar och bildar drusen (vita fläckar, Fig. 22) och tillåter läckage av vätska bakom fovea. Konerna i fovea dör orsakar central visuell förlust så att vi inte kan läsa eller se fina detaljer.
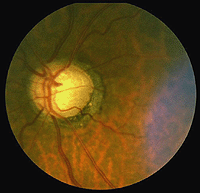
glaukom (Fig., 23) är också ett vanligt problem vid åldrande, där trycket i ögat blir förhöjt. Trycket stiger eftersom ögonets främre kammare inte kan utbyta vätska ordentligt med de normala vattenhaltiga utflödesmetoderna. Trycket i glasögonkammaren stiger och äventyrar blodkärlen i synnerven huvudet och så småningom axoner av ganglion celler så att dessa vitala celler dör. Behandling för att minska det intraokulära trycket är viktigt vid glaukom.
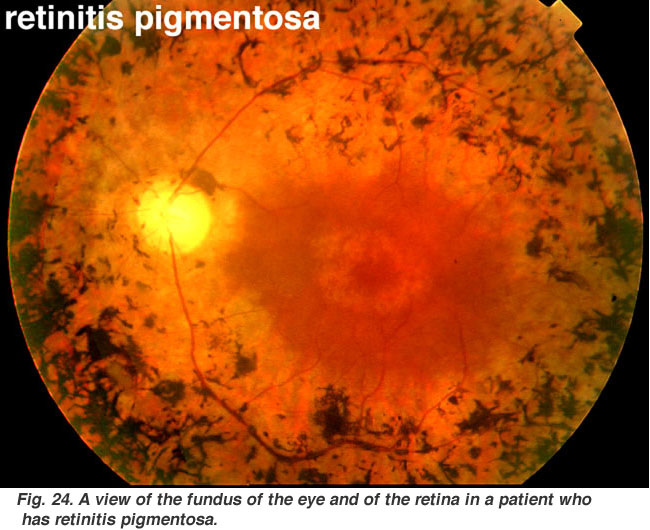
Fig. 24., En bild av ögonbotten och näthinnan hos en patient som har retinitis pigmentosa |
|
Retinits pigmentosa (Fig. 24) är en otäck ärftlig sjukdom i näthinnan för vilken det inte finns något botemedel för närvarande. Det kommer i många former och består av ett stort antal genetiska mutationer som för närvarande analyseras., De flesta av de felaktiga generna som har upptäckts gäller stångfotoreceptorerna. Stavarna i den perifera näthinnan börjar degenerera i tidiga skeden av sjukdomen. Patienterna blir nattblinda gradvis som mer och mer av den perifera näthinnan (där stavarna bor) blir skadad. Ibland reduceras patienterna till tunnelseende med endast fovea skonade sjukdomsprocessen. Karakteristisk patologi är förekomsten av svart pigment i den perifera näthinnan och förtunnade blodkärl vid synnerven huvudet (Fig. 24).,
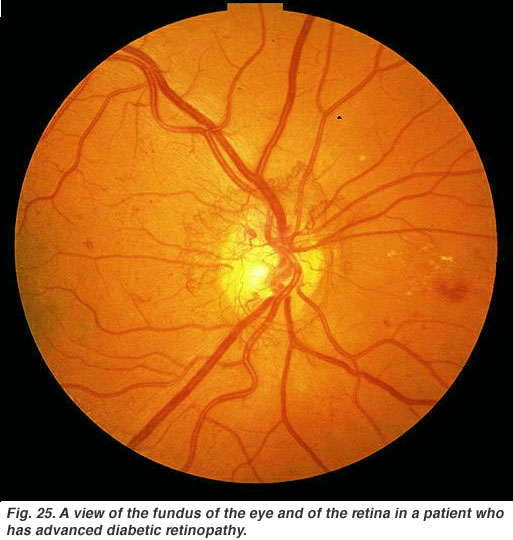
diabetisk retinopati är en bieffekt av diabetes som påverkar näthinnan och kan orsaka blindhet (Fig. 25). De vitala närande blodkärlen i ögat blir komprometterade, förvrängda och multipliceras på okontrollerbara sätt. Laserbehandling för att stoppa blodkärlsproliferation och läckage av vätska i näthinnan, är den vanligaste behandlingen för närvarande.
9. Hänvisningar.
Balashov NA, Bernstein PS. Rening och identifiering av komponenterna i de humana makulära karotenoidmetabolismvägarna. Investera Oftal Vis Sci.1998;39:s38.
Hageman GS, Johnson LV., Det fotoreceptor-retinala pigmenterade epitelet gränssnittet. I: Heckenlively JR, Arden GB, redaktörer. Principer och praxis för klinisk elektrofysiologi av syn. St. Louis: Mosby År Bok; 1991. s. 53-68.
Harrington, D.-O. och Drake, M. V. (1990) Den Visuella Fält, 6th ed. Mosby’. St. Louis.
Hayreh SS. Segmentell karaktär av den koroida vaskulaturen. Br J Oftal. 1975;59:631–648.
Kolb H. den neurala organisationen av den mänskliga näthinnan. I: Heckenlively JR, Arden GB, redaktörer. Principer och metoder för klinisk elektrofysiologi av syn. St. Louis: Mosby År Bok Inc.; 1991., s. 25-52.
Polyak SL. Näthinnan. Chicago: University of Chicago Press, 1941.
Rodieck RW. Ryggradsdjur näthinnan: principer för struktur och funktion. San Francisco: W. H. Freeman and Company, 1973.
Snodderly DM, Auran JD, Delori FC. Det makulära pigmentet. II. rumslig fördelning i primat näthinnan. Investera Oftal Vis Sci. 1984;25:674–685.
Snodderly DM, Weinhaus RS, Choi JC. Neural-vaskulära relationer i centrala näthinnan hos Makakapor (Macaca fascicularis). J Neurosci. 1992;12:1169–1193.
Van Buren JM. Det retinala ganglioncellskiktet., Springfield (IL): Charles C. Thomas; 1963.
Yamada E. vissa strukturella egenskaper hos fovea centralis i den mänskliga näthinnan. Arch Oftal. 1969;82:151–159.
Zhang HR. Scanning elektron-mikroskopisk studie av korrosion kastar på retinal och koroidal angioarchitecture hos människa och djur. Prog Ret Öga Res. 1994;13:243-270.