Gränserna i Psykologi
Inledning
Gäspningar är en ofrivillig sekvens av mun öppnas, djup inspiration, korta andningsuppehåll, och en mer eller mindre långsam utandning (Baenninger, 1997; Walusinski och Deputte, 2004; Guggisberg et al., 2010; Krestel m.fl., 2018). När det framkallas kan en gäspning inte helt undertryckas. Därför har det definierats som ett stereotypt eller reflexliknande mönster (Lehmann, 1979; Provine, 1986)., I Homo sapiens har flera hypoteser lagts fram med variabelt stöd för att förklara mekanismer och funktioner för spontan gäspning, såsom syrebildning (andningsfunktion orsakad av hypoxi), stressrelaterat beteende (orsakad av upphetsning) eller termoregulering (orsakad av hypertermi; Guggisberg et al., 2010, Massen m.fl., 2014; Gallup och Gallup, 2019). Att vara ett fysiologiskt svar kan gäspning påverkas av interna och externa faktorer som tid på dagen (Giganti och Zilli, 2011) eller intrakraniell/hjärntemperatur (Gallup och Eldakar, 2013).,
gäspningar kan vara självstyrda och / eller visas för andra (Moyaho et al., Och med 2017. Palagi et al., 2019). I mänskliga och icke-mänskliga primater, beroende på arten, när gäspning visas för andra, kan den kommunicera hot (Troisi et al., 1990; Deputte, 1994) och/eller fysiologiska och beteendemässiga förändringar (Provine et al., 1987; Leone m.fl. År 2015; Zannella et al., 2015). Hos människor är gäspning ett socialt modulerat svar eftersom det kan hämmas av faktisk—och inte virtuell-social närvaro (Gallup et al.,, 2019) och eftersom en gäspning kan utlösas av någon annans gäspning, som ett resultat av ett fenomen som kallas smittsam gäspning (Provine, 1989, 2005). Gäspningskontaktion kan framkallas även om gäspningen hörs men inte ses (Arnott et al., 2009, Massen m.fl., 2015).
I människor, deras fylogenetiskt närmast människoliknande aparter (schimpanser: Pan paniscus; dvärgschimpanser: Pan troglodytes) och Afrikanska apa Theropithecus gelada, smittsamma gäspningar är inte bara närvarande (Provine, 1986; Palagi et al., 2009; Tan et al., 2017, men se: Amici et al.,, 2014) men också socialt modulerade eftersom gäspningen är högst när vissa kategorier av individer är inblandade (t.ex. kin, gruppmedlemmar, dominanter; Palagi et al., 2009; Campbell och de Waal, 2011, 2014; Norscia och Palagi, 2011; Demuru och Palagi, 2012; Massen et al., 2012)., Två huvudargument har presenterats för att förklara denna sociala asymmetri i smittsam gäspning, som har grupperats i två huvudhypoteser: den emotionella Biashypotesen (EBH), som förbinder smittsam gäspning till känslomässig överföring och den Attentionella Biashypotesen (ABH), som anser smittsam gäspning som ett motorsvar som är föremål för skillnader i uppifrån och ner attentionella processer (Palagi et al., 2020).,
EBH förutspår att den sociala asymmetrin som observerats i gäspningar återspeglar skillnader i olika sociala bindningar, en proxy av känslomässig bindning, mellan individer. Denna hypotes stöds av bevis på att yawn contagion rates följer en empatisk trend (sensu Preston och De Waal, 2002), som är högst mellan individer som delar ett starkaste känslomässigt band. Specifikt fann Norscia och Palagi (2011) att hos människor yawn smitta är störst som svar på anhöriga och vänner än som svar på bekanta och främlingar., I vuxen schimpanser gäspa smitta är större i mellan-gruppen jämfört med ut-grupp medlemmar (Campbell och de Waal, 2011) och i bonoboernas gäspa smitta priser är störst mellan individer som affiliate mer med varandra (Demuru och Palagi, 2012). I en jämförande undersökning inklusive både människor och bonobos, Palagi et al. (2014) fann att gäspningen påverkades av sambandet mellan individer mer än av de arter som försökspersonerna tillhörde., Dessutom ökar hos människor yawn smitta med ålder när förmågan att identifiera andras känslor ökar och minskar med ålderdom när sådan förmåga minskar (Wiggers och van Lieshout, 1985; Anderson och Meno, 2003; Saxe et al., 2004; Singer, 2006; Millen och Anderson, 2011; Bartolomeus och Cirulli, 2014). Yawn smitta ökar från spädbarn till vuxen ålder även i schimpanser (Madsen och Persson, 2013).
ABH förutspår att den sociala asymmetrin som observeras vid gäspningskontaktion kan bero på skillnader i social, visuell uppmärksamhet (Massen och Gallup, 2017)., I synnerhet skulle högsta nivåer av smittsam gäspning bero på den extra uppifrån och ner, selektiv visuell uppmärksamhet åt individer som är mer relevanta för observatören,såsom bekanta ämnen, eftersom det förekommer hos människor och geladas eller dominanter, eftersom det förekommer i schimpanser eller bonobos (Yoon och Tennie, 2010; Massen et al., 2012; Massen och Gallup, 2017). Enligt Massen och Gallup (2017) skulle ABH backas upp av befintliga bevis på den olika visuella detekteringen och visuell perceptiv kodning av ansikten hos bekanta och/eller grupperade ämnen jämfört med okända (t. ex.,, Buttle och Raymond, 2003; Ganel och Gosen-Gottstein, 2004; Jackson och Raymond, 2006; Michel et al., 2006).
i denna studie analyserade vi data om gäspningar som samlats in över 9 år på människor i sina naturliga inställningar och vi extrapolerade de fall där gäspningen som emitterats av ett ämne kunde höras men inte ses av en potentiell responder (auditiv gäspning). Genom att endast överväga de fall där den visuella cue av gäspande stimulansen inte var detekterbar, verifierade vi om den sociala asymmetrin som tidigare observerats i gäspningar fortsatte eller inte., I synnerhet testade vi följande alternativa förutsägelser som härrör från de två hypoteserna som presenteras ovan (EBH och ABH).
Prediction 1a: enligt EBH påverkas graden av gäspningskontaktion av styrkan hos den interindividuella sociala bindningen—en proxy för den emotionella bindningen-i sig och inte av en annan uppifrån och ner, selektiv visuell uppmärksamhet åt vissa individer i synnerhet., Om denna hypotes stöds förväntar vi oss att observera den sociala biasen även när den visuella cueen av gäspstimulansen är utesluten och graden av hörsel smittsam gäspning är högre mellan starkt bundna jämfört med svagt bundna individer.
Prediction 1b: enligt ABH, de högre nivåerna av gäspning smitta mellan starkt bundna jämfört med svagt bundna individer skulle kopplas till den närmaste top-down, selektiv visuell uppmärksamhet som individer betalar till individer som är relevanta för dem, t. ex. familj och vänner., Om denna hypotes stöds bör den sociala bias som observeras i gäspningen försvinna när endast auditiva gäspningar beaktas eftersom den visuella cue inte kan besökas av den potentiella respondern.
material och metoder
datainsamling och operativa definitioner
för denna studie ansåg vi de vokaliserade gäspningar som emitterats av ett ämne som endast kunde höras-men inte ses-av en potentiell responder (härefter: auditiva gäspningar). Emittern och de potentiella respondenterna måste ligga inom ett intervall på ≤5 m., Vocalized yawns involverade användningen av vokalveck och de gäspningar som endast involverade tung inspiration/utgång betraktades inte som vocalized; 294 fall av auditiva yawns extrapolerades från en datauppsättning av totalt 2001 yawning bouts samlade över 9 år-från 2010 till 2019 – genom att använda alla förekomster provtagningsmetod (Altmann, 1974). Specifikt, hörsel gäspningar samlades in från November 2010 till maj 2019, från 05.30 am till 02.30 am, på mänskliga kaukasiska ämnen, i åldern 18-77 år, under deras rutinmässiga aktiviteter, t. ex. på arbetsplatser, över måltider, under sociala möten etc.,, med ämnena är omedvetna om att observeras och i avsaknad av någon uppenbar extern källa till ångest. Den auditiva gäspningsdatabasen inkluderade 193 yawner-potential responder dyads. Beroende på situationen registrerades informationen, onoterade, genom alfanumeriska koder och trädde direkt i beräkningsblad, skrivna i mobiltelefoner eller skrivna på papper och sedan in i beräkningsblad för efterföljande utarbetande. Grundläggande information som ålder och förhållandet mellan människor var känt för författarna., De potentiella respondenterna kodades som i icke-synförhållandet när huvudet roterades med 180 ° med avseende på avtryckaren eller när ett fysiskt, synblockerande hinder förekom, vilket hindrade den potentiella respondenten från att se avtryckarens ansikte och kropp. Trigger och responder var aldrig helt isolerade (t. ex. i två separata rum med stängda dörrar) från varandra., Den sociala närheten samlades på fyra nivåer: 0 = främlingar, som aldrig hade träffat tidigare; 1 = bekanta, som uteslutande delade ett indirekt förhållande baserat på ett tredje externt element, det vill säga arbetsuppgift (kollegor) eller vänner gemensamt (vänner av vänner); 2 = vänner, icke-relaterade individer som delar ett direkt förhållande som inte uteslutande är relaterat till ett tredje externt element; 3 = vanliga partners och kin (r ≥ 0.25)., Tidigare litteratur rapporterar att gäspningar kan framkallas inom 5 min efter att ha tittat på någon annans gäspning (utlösarens gäspning) (Provine, 1986), med maximalt i första minuten (Provine, 2005; Palagi et al., 2014). Litteraturen rapporterar också att från den fjärde minuten finns det en högsta sannolikhet för autokorrelation(vilket innebär att närvaron av en gäspning som utförs av ett ämne vid t0 ökar sannolikheten för att ha en annan gäspning av samma ämne vid t (0+X) där X är den ökande tidsenheten; Kapitány och Nielsen, 2017)., Därför ansåg vi att gäspensvar inträffade inom ett 3-mins tidsfönster från gäspningen som släpptes av utlösaren. För att ytterligare minska autocorrelation bias, i händelse av en kedja av gäspar som avges av avtryckaren (fler gäspar som avges i 3-min tidsfönster) vi registrerade som ett svar endast den första gäspningen utförs efter uppfattningen av den sista gäspningen. Vi kodade en gäspning som ”spontan”när inget annat ämne hade gäspat i 5 min före gäspningen., kodade som: 1 = närvaro, 0 = Frånvaro; den sociala obligationen infördes med de fyra nivåer som definieras ovan (0 = främlingar; 1 = bekanta; 2 = vänner; 3 = kin); trigger och observatörens kön märktes som: m = man, f = kvinna; åldersklasserna för avtryckaren och respondern kodades enligt följande: yo = Ungdom (18-24 år gammal); ad = vuxen (25-64 år gammal); se = senior (över 65 år gammal) (Statistik Kanada, 2009); tidsluckorna kodades enligt följande: 1 = 05:30-09:00 am; 2 = 09:01 am–12:30 pm; 3 = 12:31-16:00 pm; 4 = 16:01-19:30 pm; 5 = 19:31-23:00 pm; 6 = 23:01-02:30 (Giganti och Zilli, 2011)., Databasen (se kompletterande datablad) inkluderade 84 män, 69 kvinnor, 16 ungdomar (yo), 122 vuxna (ad) och 15 senior (se)., För att testa om förekomsten av gäspning smitta påverkades av faktorerna bond (0 = främlingar; 1 = bekanta; 2 = vänner; 3 = kin), kön av avtryckaren (Trigger_sex), kön av responder (Responder_sex), åldersklassen av avtryckaren (Trigger_ageclass), och åldersklassen av responder (Responder_ageclass), och tidslucka (från 1 till 6), använde vi en generaliserad linjär blandad modell (GLMM) som inkluderade dessa fem prediktorer som fasta effekter och triggers (Trigger) och responders (responder) identiteter som slumpmässiga effekter. Vi monterade modellerna i R (R Core Team, 2018; version 3.5.,1) Använda funktionen lmer i r-paketet lme4 (Bates et al., 2015). Vi fastställde betydelsen av den fullständiga modellen i jämförelse med en null-modell som endast omfattar de slumpmässiga effekterna (Forstmeier och Schielzeth, 2011). Vi använde ett sannolikhetsförhållande test (Dobson, 2002) för att testa denna betydelse (ANOVA med argument ”Chisq”). Vi beräknade p-värdena för de enskilda prediktörerna baserat på sannolikhetsförhållande mellan full och null-modellen med hjälp av r-funktionen ”drop1” (Barr et al., 2013). Eftersom svarsvariabeln var binär använde vi en binomial feldistribution., Vi testade om samspelet mellan könen eller åldersklasserna av utlösaren och svararen var signifikanta, men som de inte var inkluderade vi dem inte i modellen. Vi använde ett multipelkontrastpaket (multcomp) för att utföra alla parvisa jämförelser för varje bindningsnivå med Tukey-testet (Bretz et al., 2010). Vi rapporterade Bonferroni-justerade p-värden, uppskattning (Est), standardfel (SE) och z-värden.
resultat

Figur 1., Linje plot av effekten av den sociala bindningen mellan trigger och responder (X-axeln) på den genomsnittliga förekomsten av akustisk gäspning smitta (Y-axeln). Vänner och anhöriga visar betydligt högre gäspning smitta frekvenser än främlingar och bekanta (Tukey test: friends versus strangers p < 0.001; kin versus strangers p < 0.001; vänner kontra bekanta p < 0.001; kin versus bekanta p < 0.001; andra kombinationer, ns).,
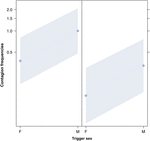
Figur 2. Linjeplot av effekten av avtryckarens kön (X-axel) på akustisk gäspningskontaktion betyder förekomst (Y-axel). Effekten av trigger sex på akustisk gäspning smitta när svararen är en kvinnlig (höger) och en manlig (vänster). Hanar gäspar framkalla mer gäspar än honor’ ones oavsett kön responder (resultat av GLMM, p = 0,022). Punkter representerar den effekt som svarsvariabeln har på den oberoende variabeln ”kön av utlösaren”, baserat på det värde som förutses av modellen., Färgade band visar 95% konfidensintervall.
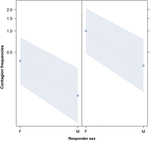
Figur 3. Line plot av effekten av kön responder (X-axeln) på akustisk gäspning smitta menar förekomst (Y-axeln). Effekt av responder kön på akustisk gäspning smitta när avtryckaren är en kvinnlig (höger) eller en manlig (vänster). Kvinnor svarar betydligt mer än män oavsett avtryckarens kön (resultat av GLMM, p = 0,021)., Poäng representerar den effekt som svarsvariabeln har på den oberoende variabeln ”svararens kön”, baserat på det värde som förutses av modellen. Färgade band visar 95% konfidensintervall.
diskussion
denna studie visar för första gången att gäspningar påverkas väsentligt av det sociala bandet mellan individer (Tabell 1) även när de utlösande stimuli är auditiva gäspningar, som vi definierade som vokaliserade gäspningar som kunde höras men inte ses (visuell cue odetekterbar, auditiv cue detekterbar)., I synnerhet är hörsel smittsam gäspning betydligt vanligare mellan anhöriga och vänner än mellan främlingar och bekanta (Figur 1). Detta fynd stödjer prognos 1a baserat på EBH och inte förutsägelse 1b baserat på ABH, vilket leder till slutsatsen att människor top-down, selektiv visuell uppmärksamhet kan inte vara den huvudsakliga drivkraften i den sociala asymmetri som observerats i gäspa smitta priser (Norscia och Palagi, 2011; Norscia et al., 2016a)., Även kön av utlösaren och kön mottagaren hade en signifikant effekt på gäspning smitta priser, med män-som triggers-besvaras av andra oftare än kvinnor (Figur 2) och kvinnor svarar oftare till andras gäspar än män (Figur 3).
i motsats till Bartholomew och Cirulli (2014) fann vi ingen ålderseffekt på gäspningar, troligen för att vår databas om auditiva gäspningar hade en stark förekomst av vuxna (25-64 år)., De högsta nivåerna av hörsel gäspning smitta hos kvinnor jämfört med män bekräftar kön bias observerats i naturalistiska förhållanden på människor som är mottagliga för gäspning smitta av Norscia et al. (2016a, b) när man överväger en större dataset som också inkluderade gäspningar som kunde ses av den potentiella respondern (med gäspande sensorisk modalitet—syn, hörsel eller båda—som inte påverkar svaret)., Genus-bias är även i partiell överenskommelse med tidigare erhållna resultat i kontrollerade inställningar, inklusive visuell (Chan och Tseng, 2017, men se Norscia och Palagi, 2011; Bartolomeus och Cirulli, 2014). Det har varit hypoteser att den höga graden av gäspning smitta hos kvinnor kan informera Emotionell smitta (Norscia et al., 2016a), mot bakgrund av enligt uppgift högre empatiska förmågor-relaterade till Moderskap—hos kvinnor jämfört med män (Christov-Moore et al., 2014)., Men denna fråga är fortfarande under debatt eftersom kulturella skillnader mellan mänskliga samhällen kan forma social bindningsdynamik på ett annat sätt. Det är därför komplicera, i detta skede av kunskap, att urskilja kulturella faktorer, inter-personlig relation kvalitet, och kön inflytande i fördelningen av gäspning smitta. Våra resultat visar också att män presterar bättre än kvinnor som triggers och den mest parsimonious hypotesen för detta kan vara att mäns vocalizations kan höras bättre i naturliga inställningar, ofta kännetecknas av bakgrundsljud., Faktum är att uppfattningen av röstkön i första hand bygger på den grundläggande frekvensen som i genomsnitt är lägre av en oktav hos män än kvinnliga röster, med lägre frekvensröstningar som reser längre än högfrekventa (Marten och Marler, 1977; Latinus och Taylor, 2012). Vi vet dock att det inte finns någon specifik studie som tar itu med den möjliga könsfördelningen i gäspningsövning och ytterligare undersökning med experimentella försök i kontrollerat tillstånd är därför nödvändig för att verifiera denna spekulation.,
i denna studie fann vi också att skillnaderna i yawn smitta priser mellan kategorier (familj och vänner, främlingar, och bekanta) inte kan förklaras av skillnader i top-down, selektiv visuell uppmärksamhet. Detta fynd är i linje med tidigare litteratur. Smittsam gäspning verkar involvera hjärnområden som är mer relaterade till orienteringsnätverket än uppifrån och ner relaterade områden . Dessutom är gäspningskontaktion varken känslig för de sensoriska ledtrådar som finns i signalen (auditiv, visuell eller audiovisuell) (Arnott et al.,, 2009; Norscia och Palagi, 2011), och inte heller påverkas av det visuella perspektivet av den utlösande stimulus (gäspar i riktlinjerna för 90°, 180° och 270° kan utlösa gäspningar svar som frontal, 0° gäspar; Provine, 1989, 1996). Chan och Tseng (2017) fann att förmågan att upptäcka en gäspning som sådan (perceptuell detekteringskänslighet) var relaterad till varaktigheten av blicken mot ögonen på det stimulansfria ansiktet, men ögonblicksmönster kunde inte modulera smittsam gäspning., I schimpanser var smittsamma gäspningsfrekvenser högsta mellan samma grupp än olika grupppersoner, även om respondenterna tittade längre på ut-grupp schimpansvideor (Campbell och De Waal, 2011). Argumentet att visuell selektiv uppmärksamhet kan fördomar gäspa smitta i en viss riktning (ämnen som svarar mer på familjen än främlingar) undergrävs också av frånvaron av något specifikt mönster av social uppmärksamhet hos människa och icke-mänsklig primat. Via ögonspårning (appliceras för att mäta visningstiden) och genom att visa okända ansikten till deras experimentella ämnen, Méary et al., (2014) observerade att människor var sneda mot egna ras ansikten medan rhesus macaques uppmärksamhet lockades mer av nya än av samma art ansikten. Kawakami et al. (2014) observerade att mänskliga ämnen ägnade större uppmärksamhet åt Etniska gruppmedlemmars ögon och näsa och mun hos etniska gruppmedlemmar. Samma studie visade också att visuell uppmärksamhet inte var beroende av målrasen. Genom att mäta hur länge försökspersonerna tittade på skärmen, Whitehouse et al., (2016) observerade att Barbary macaques ägnade mer uppmärksamhet åt att skrapa videor av icke-främling än främmande individer men noterade också att inom de icke-främlingar betalade macaques mest uppmärksamhet åt de individer som de delade en svag social bindning med. Genom att mäta glansfrekvenser observerade Schino och Sciarretta (2016) att mandrills tittade mer på sina egna släktingar än hos icke-anhöriga men också mer på dominerande än hos underordnade gruppkamrater. Därför beskriver dessa studier (som används för att stödja ABH) inget enda mönster av selektiv uppmärksamhet., En annan viktig punkt att tänka på är själva definitionen av förtrogenhet och gruppmedlemskap som antagits av de flesta studier som används för att stödja ABH (Massen och Gallup, 2017). Dessa studier visade bättre visuell upptäckt och visuell perceptiv kodning av ansikten av bekanta/gruppämnen jämfört med okända men definierade förtrogenhet och gruppmedlemskap inte på grundval av de personliga relationerna mellan individer. I stället definierades förtrogenhet eller gruppmedlemskap på grund av indirekt kunskap (t. ex., foto av kända personer eller av en person som redan visas i pre-trial faser) eller vanliga race (t ex, Buttle och Raymond, 2003; Ganel och Gosen-Gottstein, 2004; Jackson och Raymond, 2006; Michel et al., 2006). Denna definition är bra för syftet med dessa studier, men det är inte så mycket bra om resultaten används för att föreslå alternativa förklaringar för det inflytande som verklig social bindning—baserat på verkliga relationer—kan ha på ett fenomen, i detta fall gäspning smitta. Till exempel Michel et al., (2006) observerade att kaukasiska och asiatiska ämnen bättre kunde känna igen samma ras ansikten men denna skillnad var inte närvarande i asiatiska ämnen som hade bott i ungefär ett år bland kaukasier. En annan diskussionspunkt gäller förekomsten av gäspningskontaktion hos barn med autismspektrumstörning (ASD), som ofta visar förändringar i visuell uppmärksamhet (Richard och Lajiness-O ’ Neill, 2015). I ASD-barn kan gäspkontagion vara frånvarande (Senju et al., 2007), osäkra (Helt et al.,, 2010), eller liknar vanligtvis utveckla barn när ämnena induceras att omdirigera deras uppmärksamhet video stimulans under experimentella försök (Usui et al., 2013). I en nyligen genomförd studie, Mariscal et al. (2019) fann att gäspningar i ASD-barn var positivt relaterade till blodkoncentrationen av oxytocin, det hormon som är involverat i föräldra-och social anknytning (Decety et al., 2016) och ansåg att gäspningskontaktion hos ASD-barn kan relateras till varierande medelkoncentrationer av oxytocin i olika studiekohorter (Mariscal et al., 2019)., Detta fynd är i linje med EBH-hypotesen som länkar yawn smitta till social bindning, vilket kan återspegla känslomässig bindning.
slutsats
vår studie lägger till diskussionen om de mekanismer som ligger till grund för den sociala asymmetrin i gäspningskontaktion (för kritiska recensioner: se Adriaense et al., 2020; Palagi m.fl., 2020), genom att visa att gäspning smitta är förmodligen förknippad med bottom-up, snarare än med top-down, selektiv uppmärksamhet., Bottom-up-uppmärksamhet leder främst av den sensoriska uppfattningen av den eliciting stimulansen, medan top-down, selektiv uppmärksamhet är en frivillig, hållbar process där ett visst objekt väljs internt och fokuseras på eller undersöks (Katsuki och Constantinidis, 2014). I detta avseende hördes den akustiska stimulansen (auditiv gäspning) som utlöstes av utlösaren och kunde framkalla ett gäspningsrespons i mottagaren, även om mottagaren inte betalade någon frivillig visuell uppmärksamhet åt utlösaren., Dessutom var gungningsreaktionsfrekvensen Socialt modulerad, med auditiv gäspningskontaktion som högst hos individer som var starkast bundna till varandra. Därför är top-down selektiv uppmärksamhet inte den främsta drivkraften för den sociala asymmetrin som observeras vid gäspningskontaktion, vilket verkar vara ett stimulansdrivet fenomen-relaterat till bottom-up-uppmärksamhetsprocesser. Ytterligare undersökning är nödvändig för att förstå om och på vilket sätt andra former av uppmärksamhet eller pre-uppmärksam stadier kan påverka gäspning smitta.,
datatillgänglighet uttalande
datauppsättningen som används för denna studie tillhandahålls i det kompletterande materialet.
Etik Uttalande
Den studie som involverar mänskliga deltagare har granskats och godkänts av Comitato di Bioetica d’Ateneo (University Bioetiska Kommittén)—Universitetet i Turin (ref. nr 451945). Skriftligt informerat samtycke för deltagande krävdes inte för denna studie i enlighet med den nationella lagstiftningen och de institutionella kraven.
Författarbidrag
I och EP utförde datainsamling och tänkt och skrev manuskriptet., Az hjälpte till med datainsamling och manuskript revision, siffror och tabeller. MG utförde statistiska analyser och skrev den relaterade delen av manuskriptet.
intressekonflikt
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
författarna vill tacka Prof.Roberto Barbuti och Prof. Cristina Giacoma för att ha stött forskningen vid universitetet i Pisa respektive universitetet i Turin.,
Supplementary Material
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–266. doi: 10.1163/156853974×00534
PubMed Abstract | CrossRef Full Text | Google Scholar
Anderson, J. R., and Meno, P. (2003). Psychological influences on yawning in children. Curr. Psychol. Lett. 11, 1–7.
Google Scholar
Bartholomew, A. J., and Cirulli, E. T. (2014)., Individuell variation i smittsam gäspning känslighet är mycket stabil och till stor del oförklarlig av empati eller andra kända faktorer. PLoS One 9:e91773. doi: 10.1371 / tidskrift.pone.0091773
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Bretz, F., Hothorn, T., och Westfall, P. (2010). Flera Jämförelser Med R. Boca Raton, FL: Chapman & Hall.
Google Scholar
Deputte, B. L. (1994). Etologisk studie av gäspning i primater. I., Kvantitativ analys och studie av orsakssamband hos två arter av gamla världsapor (Cercocebus albigena och Macaca fascicularis). Etologi 98, 221-245. doi: 10.1111 / j. 1439-0310.1994.tb01073.X
CrossRef Full Text | Google Scholar
Dobson, A. J. (2002). En introduktion till generaliserade linjära modeller, 2nd Edn. Boca Raton, FL: Chapman and Hall.
Google Scholar
Katsuki, F. Och Constantinidis, C. (2014). Bottom-up och top-down uppmärksamhet: olika processer och överlappande neurala system., Neuroforskare 20, 509-521. doi: 10.1177/1073858413514136
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Sångerska, T. (2006). Den neuronala grunden och ontogeni av empati och sinnesläsning: granskning av litteratur och konsekvenser för framtida forskning. Neurosci. Biobehav. Rev. 30, 855-863. doi: 10.1016 / j. neubiorev.2006.06.011
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Statistics Canada (2009). Åldersgrupper, Livscykelgrupper. Ottawa: Statistik Kanada.,
Google Scholar