アセチルCoA
定義
アセチルCoAまたはアセチル補酵素Aは、生化学反応にアセチル基を追加する細胞呼吸(エネ これらの反応は、タンパク質、炭水化物、およびアデノシン三リン酸(ATP)、乳酸、およびケトン体の形でエネルギー源を提供する脂質の代謝に使用されます。 最近の研究はアセチルCoAがまた細胞内のメカニズムの重要な規定する役割を担うことを示します。 また、断食や飢えたときのエネルギー生産にも不可欠です。,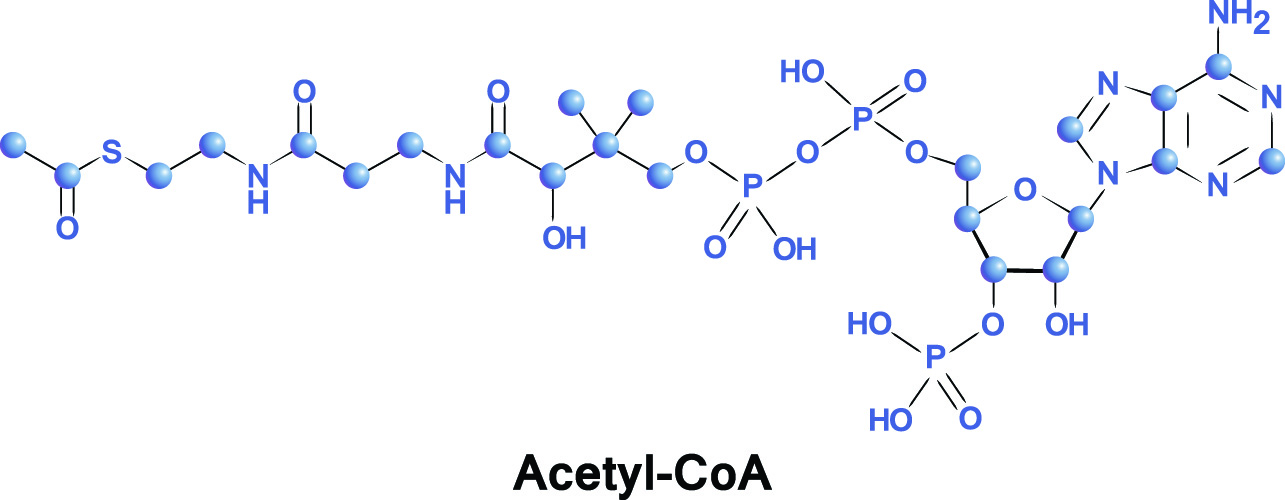
アセチルCoA形成
アセチルCoA形成は、細胞ミトコンドリアの内側または外側に発生します。 代謝産物(代謝に必要な物質)として、アセチルCoAは自由に入手できなければならない。 それは炭水化物(ブドウ糖)および脂質(脂肪酸)の異化(故障)によって作り出すことができます。 その主な仕事は、他の分子にアセチル中の炭素原子を転送することです。
アセチルco-Aの成分は、驚くことではないが、アセチルおよび補酵素Aである。アセチル基は、化学式CH3COで表される。, アセチルは、炭水化物の誘導体であるピルビン酸の分解によって生成される。 ピルビン酸塩が分解すると、小さな結合炭素分子(C2)が生成されます。 それらがCoAと反応するとき、結合された分子はアセチルCoAになります。
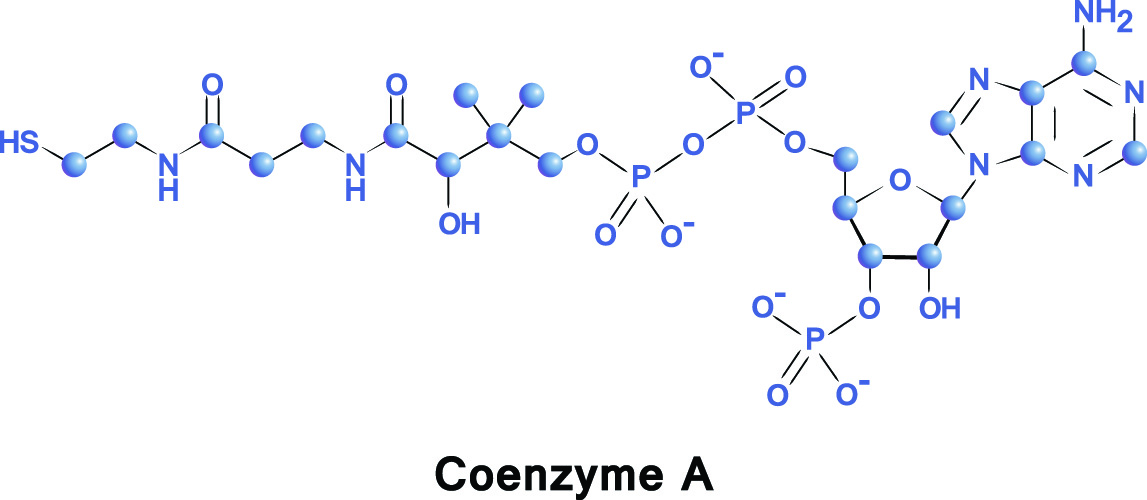
補酵素Aは補因子であり、酵素が効果をもたらすのを助ける。 Co-Aは、ビタミンB5(パントテン酸またはパントテン酸)の摂取によって生成される。, このビタミンの自然な源はキャベツおよびブロッコリー、全穀物およびポテトである。 補酵素Aの化学式はC23H38N7O17P3Sです。
多くのタイプの腸内細菌はある特定のアミノ酸からパントテン酸を製造します。 ボディのpantothenateのレベルが低いとき、CoAおよびアセチルCoAのレベルはまた低いです。 CoAの生産が他のビタミン産生経路と重複するので、これらはまたCoAおよびアセチルCoA両方の利用可能性に影響を与えることができます。 競合するビタミンの例は、葉酸およびチアミンである。,

アセチルは、制御された状況で補酵素Aと結合する。 これらの形成経路は、以下の段落により詳細に記載される。 Krebのサイクルまたはクエン酸サイクルの基本的な知識は、アセチルCoAについて学ぶときに非常に役立ちます。
グルコースを介したアセチルCoA形成
アセチルCoA形成は、最も一般的にグルコース異化の間に発生します。, 炭水化物が消化酵素によって分解された後、細胞のグルコース代謝または解糖の第一段階が始まることがあります。 解糖は、グルコース分子の分解である。 このメカニズムは、細胞サイトゾルで起こる。 下の画像では、解糖は紫色のボックスで表されています。,
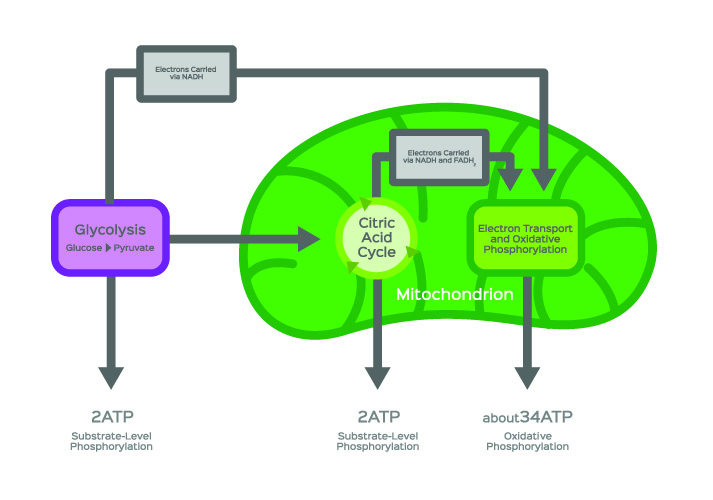
簡略化された用語では、解糖反応は、二つの水素イオン、二つのATP分子の総ゲイン、および単一のグルコース分子からcūhūoū)。
C6グルコースは二つのC3ピルビン酸分子になります。 ピルビン酸の完全な化学式はC3H3O3です–あなたはピルビン酸とグルコースの二つの化学式を見れば、グルコースはほぼ半分に分割されています。, 解糖反応の間に水素原子が放出されている。
グルコース代謝の第二段階は、酸素の有無またはそれを使用する細胞の能力に依存する。 利用可能な酸素がないかまたは限られている場合、ピルビン酸塩は、乳酸産生(嫌気性呼吸)につながる嫌気性経路を移動する。,

好気性呼吸(酸素の存在下でのエネルギー産生)は、しかし、ピルビン酸 クエン酸サイクルは、そうでなければトリカルボン酸(TCA)サイクルまたはKrebのサイクルとして知られている、細胞内エネルギー生産の基礎石です。
好気性サイクルへのエントリは、三つの準備段階が行われた後にのみ発生することができます。, まず、二つのピルビン酸分子(C3)は酸化的リン酸化(電子交換)を受ける。 この工程は、アセチルCoAを含まない。
第二に、エネルギー放出相はADPを四つのATP分子に変換する。 再び、アセチルCoAは必要とされない。
第三段階は、ピルビン酸のアセチルへの変換であり、その後のアセチルと利用可能な補酵素Aとの結合である。,
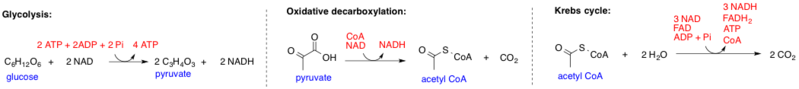
ピルビン酸のアセチルCoAへの変換は、ピルビン酸酸化脱炭酸と呼ばれる三段階のプロセスでもある。 この経路は細胞のミトコンドリアの中で起こり、ピルビン酸塩の分子は活動的な輸送によってmitochondriaに入ります。
まず、負に帯電したカルボキシレートアニオン基(COO−)をピルビン酸(C3H4O3)から酵素ピルビン酸デヒドロゲナーゼによって除去し、二酸化炭素(CO2)を形成する。 ピルビン酸は現在C2H3Oまたはアセチルになっています。,
第二に、カルボキシレートアニオン基の負電荷は補因子反応(NAD+およびNADH反応)に役立ちます。 Krebのサイクルに精通している場合は、これら二つの補因子がエネルギー生産において非常に重要な役割を果たすことを知っているでしょう。
ピルビン酸酸化脱炭酸の最終ステップは、補酵素Aのアセチルへの結合である。 この高エネルギーで非常に反応性の高い結合は、アセチル基と補酵素Aの硫黄との間に形成され、アセチル-CoAを形成する。 この分子は現在、クエン酸サイクルに直接contributeすることができます。,
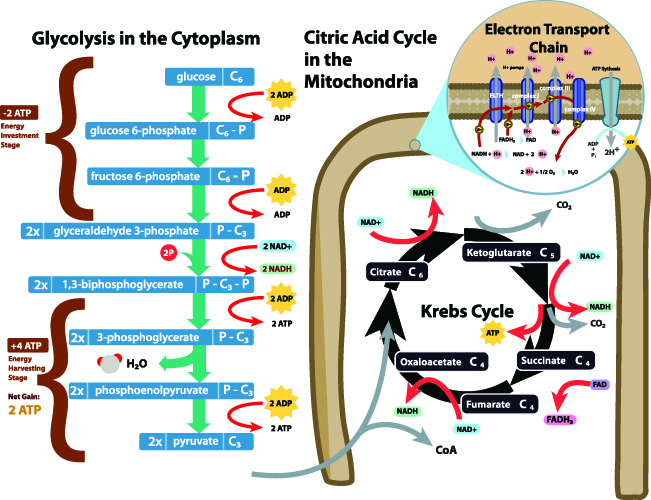
クエン酸サイクルは常に補酵素AとアセチルCoAを形成し、再生します。 アセチルCoAの単一の分子はATPの10から12分子を作り出します。 アセチル基がアセチルCoAから放出された場合、残りの補酵素Aは、クエン酸サイクルに再入る前にピルビン酸のアセチルCoAへの変換を助ける。,
脂肪酸によるアセチルCoA形成
アセチルCoA形成はまた、脂肪酸異化作用によって起こると言われているが、このアセチルCoAは糖代謝の産物であることが理解されている。 アセチルCoAが脂質に変えることができる逆もまた同じであるので別の役割と時々混乱されます;その本当の役割は単糖類(ブドウ糖)の新陳代謝の触媒としてあります。
脂肪代謝において、摂取されたトリグリセリドは、それらの最小の形態を含まない脂肪酸に分解され、これらは血流に輸送される。, 脂肪組織の脂肪細胞(adipocytes)はグリセロールが付いているこれらの脂肪酸を結合し、バックアップエネルギー源として役立つためにトリグリセリドの鎖とし Carbohydrates化物源が低い場合、脂肪からエネルギーを得ることができる。
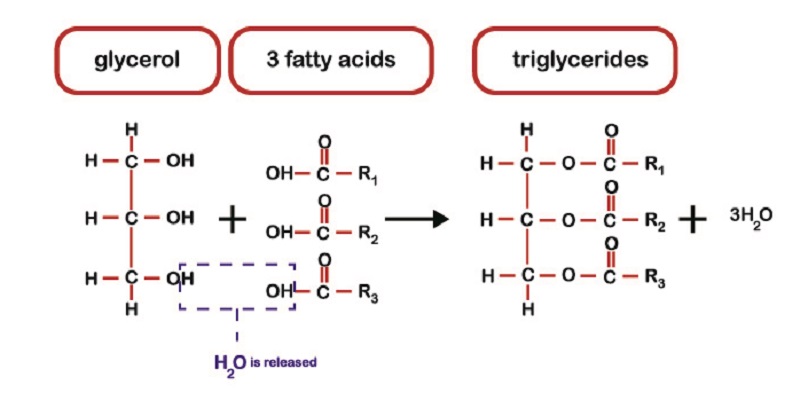
これは、負に帯電した脂肪酸とグリセロールへのトリグリセリドの脂肪分解 脂肪酸の酸化(酸素を加える)反応は、アセチルCoAではなく脂肪アシルCoAを形成する。 これは混乱があるところである。, 体はKrebのサイクルでアシルCoAを使用することはできません。 それはアセチルCoAに変換されなければならない。
脂肪アシルCoAのアセチルCoAへの変換は、ミトコンドリア内で起こり、酵素アシルCoAデヒドロゲナーゼおよび脂肪酸鎖中のすべての炭素がアセチルCoA分子に変換されるまで続く一連の反応を必要とする。 これらはKrebのサイクルに入ることができます。 脂肪酸からのATPの収穫は単糖類のそれより大いに低い–ちょうど14から炭水化物の36。,
アセチルCoA構造
アセチルCoA構造は、輸送補酵素基と結合したアセチル基からなる。 補酵素は生物的分子の範囲の故障の酵素を助けます。
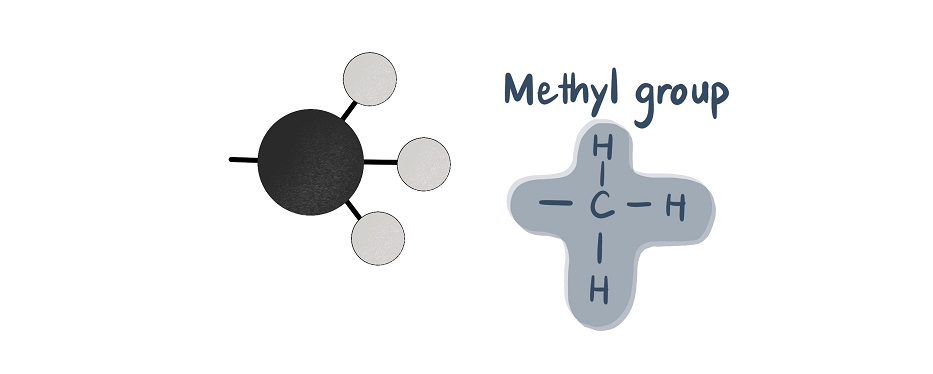
アセチル基は二つの炭素単位を含み、化学式C2H3Oを有し、二重結合を介して結合したメチル基(CH3)と二重結合カルボニル基(CO)からなる。,
アセチルCoAにおいて、アセチル基は補酵素Aに結合する。(必須ビタミン)、リン酸、およびアデノシン二リン酸(adp)。 補酵素の部分はアセチルグループのための運送者です。 これは、適切な場所にアセチル基をもたらし、アセチル基は、クエン酸サイクル内の他の物質に二つの炭素原子を転送することができます。,
糖新生におけるアセチルCoA
糖新生は、簡単に言えば、逆の解糖である。 グルコースのレベルが低い場合、例えば糖尿病性血糖降下期または飢餓または長期断食中など、体は非炭水化物源からグルコースを作ることができる。 アセチルCoAは糖新生において重要な調節的役割を果たしている。 ほとんどの糖新生は肝臓の細胞で起こり、腎臓の細胞では軽度の反応が起こる。,

糖新生において、ピルビン酸はまずいくつかの酵素の影響下でホスホエノールピルビン酸(PEP)酸に変換されなければならない。 ピルビン酸カルボキシラーゼ-それは直接このステップに関与する多くの酵素の一つを制御するようにアセチルCoAは、この転化率を調節します。
エネルギーのためのボディの必要性および供給に関するフィードバックはまたアセチルCoA供給によって提供されます。, アセチルCoAのレベルが高いとき、ピルビン酸塩はクエン酸周期から取除かれ、貯えられます。
次のステップは、(肝臓)細胞の小胞体内のフルクトースの形態のグルコースへの変換である。 このブドウ糖は付加的で、費用効果が大きいエネルギーを提供し、またレバ 準備手順を行されたのAcetyl-CoA形成糖ます。
アセチル補酵素A:追加の役割
アセチルCoAは、多くの追加の役割を持っています。, これらの脂質、コレステロール値、およびステロイド合成の源泉となる胆汁酸塩、性ホルモン,アルドステロン、コルチゾール. これらの化学薬品およびホルモンは消化が良い、生殖の、および神経系機能の広い範囲を支える。

ケトン体、減量のフォーラムで議論の人気のあるトピックは、飢餓イベントの結果です。, オキサロ酢酸の可用性は、クエン酸サイクル内で重要であり、直接アセチルCoAの可用性に関連付けられています。 クエン酸サイクルでは、アセチルCoAはオキサロ酢酸と結合してクエン酸を形成する。
飢餓モードまたは低血糖の期間中、グリコーゲン貯蔵量が枯渇するか、または使用することができない場合。 糖新生–脂肪およびタンパク質からのグルコース合成-が必要である。 オキサロ酢酸が不足している場合、アセチルCoAは代わりにケトン体(ケトン生成)を形成する。 ケトン体では、オキサロ酢酸は必要ありません。,

糖尿病性ケトアシドーシスに苦しむ人々の呼吸で検出できるケトン体です。 ケトン体は、グルコースレベルが低いときに最も重要な器官(心臓、腎臓、および脳)にエネルギーを提供することができます。,
この非グルコースエネルギー源の使用は、非常に低から非炭水化物アトキンスダイエット(長年にわたって多くの論争を引き起こしている)や、最近では炭水化物を許可するが12-72時間の断食状態を伴う断続的な断食ライフスタイルなどの低炭水化物ダイエットの基礎でもある。 断続的な断食の長期的な影響はまだ証明されていませんが、これまでのところ結果は肯定的なようです。 炭水化物の食事療法への低速は矛盾した証拠を提供しないようである。, これらの食事のいずれかを検討している人は、最初に医師に相談し、六ヶ月の血液検査を手配する必要があります。
